Кірка покривна тканина. покривні тканини
Вони виконують роль прикордонного бар'єру, відокремлюючи нижче лежачі тканини від навколишнього середовища. Первинні покриви рослини складаються тільки з живих клітин. Вторинні і третинні покриви - в основному з мертвих з товстими клітинними стінками.
Основні функції покривних тканин:
· Захист рослини від висихання;
· Захист від потрапляння шкідливих мікроорганізмів;
· Захист від сонячних опіків;
· Захист від механічних пошкоджень;
· Регуляція обміну речовин між рослиною і навколишнім середовищем;
· Сприйняття роздратування.
Первинна покривна тканина - епідерміс, епідерміс. Складається з живих клітин. Утворюється з апікальних меристем. Покриває молоді зростаючі стебла і листя.
Епідерма сформувалася у рослин в зв'язку з виходом з водного середовища проживання на сушу з метою запобігання від висихання. Крім продихів, всі клітини епідерми щільно з'єднані між собою. Зовнішні стіни основних клітин товщі інших. Вся поверхня покрита шаром Кутіна і рослинних восків. Цей шар називається кутикулою (шкірка). Вона відсутня на зростаючих коренях і підводних частинах рослин. При пересиханні проникність кутикули значно послаблюється.
Крім основних клітин, в епідермісі є і інші, зокрема волоски, або трихоми. Вони бувають одноклітинними і багатоклітинними (Рис.2).Функціонально вони збільшують поверхню епідерми, наприклад, в зоні росту кореня, служити механічним захистом, чіплятися за опору, зменшувати втрати води. Ряд рослин мають залізисті волоски, наприклад, кропива.
Тільки у вищих рослин в епідермісі є продихи, які регулюють обмін води і газів. Якщо кутикули немає, то і відсутня потреба в продихах. Устячка - це група клітин, що утворюють устьічнимі апарат, який складається з двох замикаючих клітин і прилеглих до них клітин епідерми - побічних клітин. Вони відрізняються від основних епідермальних клітин (рис.3
). Замикають клітини відрізняються від тих, що оточують їх клітин формою і присутністю великої кількості мітохондрій та нерівномірно потовщеними стінками. Ті, які знаходяться один напроти одного, товщі інших (Рис.4)
Між замикаючими клітинами утворюється устьичная щілину, яка веде в подустьічное простір, який називається подустьічной порожниною. Замикають клітини мають високу фотосинтетичної активністю. У них міститься велика кількість запасного крохмалю і численні мітохондрії.
Число і розподіл устьиц, типи устьічних апаратів широко варіює у різних рослин. Продихи у сучасних мохоподібних відсутні. Фотосинтез у них здійснює гаметофітное покоління, а спорофіти до самостійного існування не здатні.
Зазвичай продихи розташовуються на нижньому боці аркуша. У плаваючих на водній поверхні рослин - на верхній поверхнос. У листя злаків продихи часто розташовуються рівномірно по обидва боки. Такі листи висвітлюються порівняно рівномірно. На 1мм2 поверхні може розташовуватися від 100 до 700 продихів.
Вторинна покривна тканина (перидерма). Ця тканина приходить на зміну епідермі, коли зелений колір однорічних пагонів змінюється коричневим. Вона багатошарова і складається з центрального шару камбіальних клітин - феллогена. Клітини феллогена, ділячись, назовні відкладають шар феллеми, а всередину - феллодерми (Рис.5).
Феллема, або пробка. Спочатку складається з живих тонкостінних клітин. Згодом їх стінки просочуються суберином і рослинними воском і відмирають. Вміст клітини наповнюється повітрям.
Функції феллеми:
· Запобігає втраті вологи;
· Захищає рослину від механічних пошкоджень;
· Захищає від хвороботворних мікроорганізмів;
· Забезпечує термоізоляцію, так як клітини заповнені повітрям.
Клітини феллогена, розташованого в самій епідермі, що підлягає субепідермальной шарі, рідше - в глибоких шарах первинної кори, є генеруючої основою первинної кори.
Статті і публікації:
Історична довідка
Перші роботи по виділенню і вивчення білкових препаратів були виконані ще в 18 ст., Проте в той період дослідження білків носили описовий характер. На початку 19 ст. були зроблені перші аналізи елементного складу білків (Ж. Л. Гей-Лю ...
Мікробіологічний контроль виробництва кисломолочних продуктів
Завдання мікробіологічного контролю зводяться до забезпечення належної спрямованості мікробіологічних процесів і дотримання санітарно-гігієнічних умов виробництва. Виходячи з цього, санітарно-мікробіологічний контроль, виробниц ...
Вчення Павлова про дві сигнальні системи дійсності. Вікові особливості розвитку і взаємодії першої та другої сигнальної системи. Роль функціональної асиметрії мозку в розвитку мови
Сигнальні системи - це системи нервових процесів, тимчасових зв'язків і реакцій, що формуються в головному мозку в результаті впливу зовнішніх і внутрішніх подразнень і забезпечують тонке пристосування організму до навколишнього середовища. ...
Ботаніка-наука про рослини.
Отліч.черти раст.клеткі
Пластида
3.Тканямі
Освітні тканини.
Ботаніка-наука про рослини.
Ботаніка охоплює широке коло проблем: закономірності зовнішнього і внутрішньої будови(Морфологія і анатомія) рослин, їх систематику, розвиток протягом геологічного часу (еволюція) і родинні зв'язки (філогенія), особливості минулого і сучасного поширення по земної поверхні(Географія рослин), взаємини з середовищем (екологія рослин), складання рослинного покриву (фітоценологія, або геоботаніка), можливості та шляхи господарського використання рослин (ботанічне ресурсоведеніе, або економічна ботаніка).
По об'єктах дослідження в ботаніці виділяють фікологов (альгологів) - науку про водорості, мікологія - про грибах, ліхенологія - про лишайники, бріологія - про мохи та ін .; вивчення мікроскопічних організмів, переважно зі світу рослин (бактерій, актиноміцетів, деяких грибів і водоростей), виділяють в особливу науку - мікробіологію. Хворобами рослин, що викликаються вірусами, бактеріями і грибами, займається фітопатологія.
Ботаніка користується як спостереженням, так і порівняльним, історичним і експериментальним методами, що включають збір і складання колекцій, спостереження в природі і на дослідних ділянках, експеримент в природі і в умовах спеціалізованих лабораторій, математичну обробкуотриманої інформації. Поряд з класичними методамиреєстрації тих чи інших ознак досліджуваних рослин використовується весь арсенал сучасних хімічних, фізичних і кібернетичних методів дослідження.
Отліч.черти раст.клеткі
Природа і основні функції речовин різні. Найголовніші з цих речовин: прості білки; деякі вуглеводи, зокрема глюкоза, сахароза та крохмаль або близький до нього інулін, а також целюлоза, запасні жири і жироподібні речовини - сполуки первинного метаболізму; продукти вторинного метаболізму - танніди, поліфенольні сполуки, алкалоїди, ізопренпроізводние і др.относітся також звичайний у багатьох рослинах оксалат кальцію Майже всі речовини незалежно від їх природи в тій чи іншій мірі можуть знову втягуватися в процеси активного метаболізму клеткі.Важнейшая група - допоміжні матеріали. Це білки, вуглеводи (виключаючи целюлозу) і жіри.Целлюлоза і близькі до неї за хімічною природою геміцелюлози і пектинові речовини - структурні компоненти клітинної стінки. Вторинні метаболіти виконують різні функції, але найголовніша з них пов'язана з "організацією" захисту від поїдання рослин тваринами, впровадження хвороботворних мікроорганізмів і т.д. Таким чином, вони є групою захисних речовин. Оксалат кальцію найчастіше розглядається в якості кінцевого продукту метаболізму, форми "поховання" непотрібних або шкідливих клітці речовин. У техніці, харчовій промисловості та медицині широко використовуються крохмаль, глюкоза і сахароза. Танніди, або дубильні речовини, сполуки поліфенольні і ізопренпроізводние - основа для отримання більшості медичних препаратів рослинного походження. Пектинові речовини широко застосовуються в кондитерській промисловості, а також в медицині. Деякі з ергастіческіх речовин дуже отруйні. Найчастіше це алкалоїди, деякі глікозиди, поліпептиди (у блідої поганки). Значення допоміжних матеріалів для фармації і медицини.
Крохмаль, в медицині і фармації для приготування присипок, паст (густих мазей), а також при виробництві таблеток. Целюлоза, велике і промислове значенняцелюлози - з цієї речовини виготовляють бавовняні тканини і бумагу.Клеткі видільних тканин за формою зазвичай паренхімні і тонкостінні. Вони довго залишаються живими, виділяючи секрет. Клітини-ідіобласти в міру накопичення великої кількості секрету позбавляються протопласта і стінки їх нерідко опробковевают.Ткані внутрішньої секреції можуть бути представлені окремими клітинами-ідіобласти, вместилищами виділень, смоляними ходами, Ефіро каналами і молочні судини. Пластида- безбарвні або забарвлені тільця в протоплазмі рослинних клітин, що представляють собою складну систему внутрішніх мембран і виконують різні функції. Безбарвні пластиди називають лейкопластах, по-різному забарвлені - хромопласти, зелені - хлоропластами.
Тільки в рослинних клітинах присутні пластиди. Ці мембранні органели в залежності від забарвлення можна розділити на лейкопласт, хромопласти і хлоропласти. У клітці вищих рослин міститься близько 40 хлоропластів в яких відбувається фотосинтез. Як і мітохондрії, хлоропласта містять власну ДНК і рибосоми. Вони здатні до автономного розмноження, який не залежить від поділу клітини. Залежно від забарвлення, пов'язаної з наявністю або відсутністю тих чи інших пігментів, розрізняють три основних типи пластид:
Хлоропласти (зеленого кольору), - хромопласти (жовтого, оранжевого або червоного кольору) і - лейкопласт (безбарвні).
Найбільші з них - хлоропласти - досягають у вищих рослин 4-10 мкм довжини і добре помітні в світловий мікроскоп. Хлоропласти зустрічаються у всіх зелених органах рослин, лейкопласт дуже звичні в клітинах органів, прихованих від сонячного світла, - корінні, кореневищах, бульбах, а також в сітовідних елементах деяких покритонасінних. Хромопласти містяться в клітинах пелюсток багатьох рослин, зрілих пофарбованих плодах (томати, шипшина, горобина), іноді - в коренеплодах (морква).
Будова пластид може бути розглянуто на прикладі хлоропластів. Вони мають оболонку, утворену двома мембранами: зовнішньої і внутрішньої. Внутрішня мембрана вдається в порожнину хлоропласта нечисленними виростами. Мембранна оболонка відмежовує від гіалоплазми клітини матрикс хлоропласта, так звану строму. Як строма, так і вирости внутрішньої мембрани формують в порожнині хлоропласта складну систему мембранних поверхонь, що відмежовують особливі плоскі мішки, звані тилакоїди, або ламелами. Групи дисковидних тилакоидов пов'язані один з одним таким чином, що їх порожнини виявляються безперервними. Ці тилакоїди утворюють стопки (на зразок стопки монет), або грани. Тилакоїди строми об'єднують грани між собою. У мембранах тилакоїдів зосереджений найголовніший пігмент зелених рослин - хлорофіл і допоміжні пігменти - каротиноїди. Внутрішня структура хромопластов і лейкопластов простіше. Грани в них відсутні.
В стромі хлоропластів містяться ферменти і рибосоми, що відрізняються від рибосом цитоплазми меншими розмірами. Часто є один або кілька невеликих зерен первинного асиміляційного крохмалю. Генетичний апарат хлоропластів автономний, вони містять свою власну ДНК.
Основна функція хлоропластів - фотосинтез. Центральна роль в цьому процесі належить хлорофілу, точніше - кільком його модифікацій. Світлові реакції фотосинтезу здійснюються переважно в гранах, темнові - в стромі хлоропласта. І хлоропласти, і мітохондрії здатні синтезувати власні білкові молекули, так як володіють власною ДНК.
Крім фотосинтезу, в хлоропластах здійснюється синтез АТФ і АДФ (фосфорилювання), синтез і гідроліз ліпідів, асиміляційного крохмалю і білків, відкладаються в стромі.
У лейкопластах пігменти відсутні, але тут може здійснюватися синтез і накопичення запасних поживних речовин, в першу чергу крохмалю, іноді білків, рідко жирів. Дуже часто в лейкопластах формуються зерна вторинного запасного крохмалю.
Червона або оранжеве забарвлення хромопластов пов'язана з присутністю в них каротиноїдів. Вважається, що хромопласти - кінцевий етап у розвитку пластид, інакше кажучи, це старіючі хлоропласти і лейкопласт. Наявність хромопластов частково визначає яскраве забарвлення багатьох квіток, плодів і осіннього листя.
3.Тканяміназиваються групи клітин, які мають однакове походження, схожу будову і виконують в організмі однакову функцію.
Кожен тип тканини відрізняється характерним будовою клітин, строго відповідним тієї функції, яку дана тканина виконує.
Відповідно до форми, будовою і характером з'єднання клітин тканини можуть бути пухкі і щільні, живі і мертві, паренхімні і прозенхімних. Все розмаїття рослинних тканин прийнято поділяти на 6 основних груп: 1) освітні, або меристеми, 2) основні, 3) покривні, 4) механічні, 5) проводять і 6) видільні. Освітні тканини.Меристеми або освітні тканини, мають здатність до поділу і утворення нових клітин. За рахунок меристем формуються всі інші тканини і здійснюється тривалий ріст рослини. У тварин меристеми відсутні, чим пояснюється обмежений період їхнього росту. Клітини меристем відрізняються високою метаболічною активністю. Одні клітини меристем, що отримали назву ініціальних, затримуються на ембріональній стадії розвитку протягом усього життя рослини, інші поступово диференціюються і перетворюються в клітини різних постійних тканин.
Первинні меристеми здатні до поділу спочатку. У ряді випадків здатність до активного поділу може знову виникнути і у клітин, вже майже втратили цю властивість. Такі "знову" виникли меристеми називають вторинними.
У тілі рослини меристеми займають різне положення, що дозволяє їх класифікувати. Відповідно до положення в рослині виділяють верхівкові, або апікальні, бічні, або латеральні, і інтеркалярние меристеми.
Апікальні меристеми розташовуються на верхівках осьових органів рослини і забезпечують зростання тіла в довжину, а латеральні - переважно зростання в товщину. Кожен втечу і корінь, а також зародковий корінець, почечка зародка мають апікальної меристем. Апікальні меристеми первинні і утворюють конуси наростання кореня і втечі.
Латеральні меристеми розташовуються паралельно бічних поверхнях осьових органів, утворюючи свого роду циліндри, на поперечних зрізах мають вигляд кілець. Частина з них відноситься до первинних. Первинними меристема є прокамбій і перицикл, вторинними - камбій і феллоген.
Інтеркалярние, або вставні, меристеми частіше первинними і зберігаються у вигляді окремих ділянок в зонах активного росту (наприклад, у підстав междоузлии, в підставах черешків листя).
покривні тканини, Класифікація, функції.
Вони оберігають органи рослини від висихання, від температурних впливів, механічних пошкоджень та інших несприятливих впливів навколишнього середовища. При змінах віку органів і їх функцій, покривні тканини закономірно змінюють одна одну. За походженням розрізняють первинні (епідерміс, ризодерма, веламен), вторинні (перидерма) і третинні (кірка або рітідом) покривні тканини.
Первинні покривні тканини Епідерма утворюється з поверхневого шару апікальної меристеми - протодерми. Вона покриває листя, плоди, частини квітки і молоді стебла. Крім захисної функції, епідерміс регулює процеси транспірації і газообміну, бере участь в синтезі різних речовинта ін. До складу епідерми входить кілька морфологічно різних клітин: основні клітини епідерми, що замикають і побічні клітини продихів, трихоми (вирости епідерми). Клітини епідерми живі, мають ядра, лейкопласт, вакуолі, хлоропласти (тільки в замикаючих клітинах продихів). Епідерма у більшості рослин одношарова, рідше багатошарова. Клітини первинної покривної тканини щільно прилягають один до одного, і не мають межклетников. Із зовнішнього боку вся епідерміс покрита суцільним шаром кутикули (переривається тільки над устьічнимі щілинами).
Ризодерма (епіблема) утворена апикальной меристемою кореня. Вона покриває молоді кореневі закінчення і саме через ризодерма відбувається поглинання води і мінеральних солей з грунту. Крім того, вона взаємодіє з мікроорганізмами грунту, з кореня в грунт виділяються речовини, що допомагають грунтовому харчуванню. Клітини ризодерма мають дуже тонкі оболонки. У первинної покривної тканини кореня немає кутикули, внаслідок чого ці клітини мають оболонки легко проникні для води. На невеликій відстані від кінчика кореня утворюються кореневі волоски - вирости ризодерма.
Веламен, як і ризодерма, відбувається з поверхневого шару апікальної меристеми кореня. Ця своєрідна тканина покриває коріння епіфітів і деяких інших рослин, пристосованих до життя на періодично пересихаючих ґрунтах (аспідістра, аспарагус, алое, кливия). Веламен від ризодерма відрізняється багатошаровістю. Протопласт Велам відмирає і тому всмоктує воду не осмотическим, а капілярним шляхом.
Емергенци(Нім., Однина Emergenz, від лат. Emerge - з'являюся, виходжу), вирости на поверхні стебел і листя рослин, утворені не тільки епідермісом, а й глибше лежать тканинами (на відміну від волосків). Це - пекучі волоски кропиви, шипи на стеблах троянди і на плодах дурману, залізяки на листках росички і ін. Вторинна покривна тканина - Перидерма.перидермаскладна, багатошарова вторинна покривна тканина стебел і коренів багаторічних (рідше однорічних) рослин 1. Перидерма змінює первинні покривні тканини осьових органів, які поступово відмирають і злущуються. У більшості дводольних і голонасінних перидерма зазвичай з'являється протягом першого вегетативного періоду в тих частинах кореня і втечі, які припинили зростання в довжину. В основі перідерми стебла лежить вторинна меристема - феллоген.Феллоген стебла частіше виникає з клітин основної паренхіми, що лежить під епідермою і зберегла слабку мерістематічеськую активність. У коренів основою освіти феллогена може бути перицикл.Феллоген може закладатися окремими ділянками, пізніше зливаються, або відразу по всьому колу осьового органу. Перш за все утворюється ініціальний ряд сплощених за формою таблитчатих клітин. Пізніше ініціальні клітини периклінальних діляться, формуючи два ряди меристематичних клітин. Внутрішній ряд диференціюється як феллодерми(Живі паренхім-ні клітини) і далі не ділиться. Зовнішній - в ході серії периклінальних поділів формує різну кількість шарів клітин пробки, тривалий час, зберігаючи мерістематічеськую активність. Феллоген здатний ділитися також і антиклінальні, забезпечуючи правильне співвідношення тканин в розростаються органах. Пробка, феллоген і феллодерми утворюють перидерму. У міру того як формується перидерма, зелений колір пагонів переходить в бурий. «Доспілі» до осені пагони першого року здатні витримати зимові морози. пробкаскладається з відмерлих клітин, позбавлених межклетников. Їх оболонки просочені суберином. Клітини пробки повітро-і водонепроникні. Багатошарова пробка утворює захисний футляр, що оберігає живі тканини від втрати вологи, від різких температурних коливань і проникнення хвороботворних мікроорганізмів. На стовбурах і гілках деяких дерев (корковий дуб, бархат амурський) формується потужний шар пробки, що досягає декількох сантиметрів товщини. Таку пробку використовують для промислового отримання укупорочних матеріалів. Живі тканини, що лежать під пробкою, потребують газообмене і видаленні надлишку вологи. Тому в перидерма досить рано утворюються чечевички- отвори, «заповнені» пухкої виконує тканиною, що складається з закруглених паренхімних слабоопробковевшіх клітин з численними межклетниками. Через чечевички здійснюється «провітрювання» стебла. Чечевички, хто має вигляд невеликих горбків різної форми, добре помітні на поверхні молодих пагонів дерев і чагарників. Чечевички у вигляді дрібних крапин помітні і на деяких плодах, наприклад яблуках і грушах. У міру потовщення пагонів форма чечевичек змінюється. На стовбурах берези їх залишки помітні у вигляді характерних поперечних смужок і рисок. Перидерма утворюється, як сказано, у дводольних і хвойних рослин. У однодольних (пальми, драцени, алое) іноді шляхом багаторазового поділу і опробковенія периферичних клітин первинної кори утворюється захисна тканина функціонально подібна до перидермой. У багаторічних осьових органах дводольних і хвойних рослин зазвичай розвивається кілька перидерм. Кожна наступна закладається глибше попередньої. Згодом живі клітини зовнішніх перидерм і розташованих між ними тканин відмирають, утворюючи потужний покривний комплекс - корку.Она формується на стовбурах, багаторічних гілках і корінні дерев. На відміну від більш-менш гладкою перідерми кірка нерідко тріщинувата і нерівна (луската пробка). Існує, правда, і щодо гладка, так звана кільцева кірка (наприклад, у винограду). Ці відмінності пов'язані з особливостями закладення внутрішніх перидерм. Товста кірка надійно оберігає стовбури дерев від механічних пошкоджень, лісових низових пожеж, різкої зміни температур, опіків прямими сонячними променями, проникнення патогенних мікроорганізмів. Зазвичай у деревних порід кірка починає закладатися в зрілому віці: у сосни в 8-10 років, у дуба в 25-30 років. Щорічно нарощуючи, з роками вона може досягати значної товщини.
Лекція 5.
покривні тканини
епідерма
перидерма
Пробка (феллема)
Рітідом
механічні тканини
Коленхіма
склеренхіма
склереїди
До цієї категорії відносяться тканини, що покривають органи рослин. Вони захищають внутрішні тканини від шкідливих зовнішніх впливів і послаблюють дію факторів зовнішнього середовища, які при високій інтенсивності можуть бути небезпечними для рослин.
Особливо велике значення покривні тканини мають для надземних органів, що мешкають в мінливому (динамічною) повітряному середовищі.
Покривні тканини оберігають органи рослини від різких температурних коливань (сильного нагрівання або охолодження), від надмірної втрати води, шляхом випаровування, від пошкодження краплями дощу, граду, твердими частинками атмосферного пилу, від проникнення всередину рослини хвороботворних організмів. Багато з цих моментів мають значення і щодо підземних і підводних органів.
У коренів роль первинної захисної покривної тканини грає екзодерма , А у стебел і листя ≈ епідерміс (Шкірка).
На зміну первинної покривної тканини у багатьох рослин утворюється вторинна покривна тканина ≈ пробка (Феллема) , Вона є частиною комплексу тканин, званого перидермой .
Ще пізніше, в багатьох випадках утворюється досить складний тканинний комплекс ≈ кірка (Третинна покривна тканина).
Розглянемо все по порядку.
епідерма
Покривні тканини не можуть наглухо ізолювати рослину від зовнішнього середовища, рослина знаходиться в стані безперервного обміну з середовищем. Тому другий, не менш важливою, ніж захисна, функцією епідерми є регуляція газообміну і транспірації (природного випаровування води живими тканинами).
В процесі еволюції епідерміс виникла дуже давно, на самому початку пристосування рослин до умов життя на суші. Без неї не мислимо існування вищих сухопутних рослин. Вже у рініі була цілком розвинена епідерміс.
Крім типових функцій, характерних для покривної тканини, епідерміс може функціонувати як усмоктувальна тканина, бере участь в синтезі різних речовин, у сприйнятті подразнень, в русі листя. Таким чином, епідерміс багатофункціональна тканину.
У структурному відношенні епідерміс - складна тканина, оскільки до її складу входить ряд морфологічно різних елементів:
Основна тканина епідерми складається з живих щільно зімкнутих клітин, що мають нерідко звивисті стінки. За рахунок звивистості стінок збільшується сила зчеплення клітин і додатково підвищується міцність тканини.
Зазвичай основні клітини епідерми прозорі і не містять хлоропластів, а якщо хлоропласти і є, то в дуже незначній кількості. Через прозорі клітини основної тканини безперешкодно проходять сонячні промені.
Оболонки клітин шкірки потовщені нерівномірно: в кожній клітині найбільш товста зовнішня стінка, бічні стінки трохи тонше, внутрішні ще більш тонкі.
Клітини епідерми зазвичай покриті тонкою плівкою ≈ кутикулою . Вона являє собою продукт життєдіяльності цитоплазми, клітини, яка виділяє через оболонку на її поверхню рідкий кутин, твердіє в плівку.
Зазвичай кутикула неоднорідна у своїй будові і багатошарова. Кутикулярного шари нерідко просякнуті воском, що залягає у вигляді включень і прошарків.
Відкладення воску на поверхні кутикули різноманітні і мають вигляд: а) дрібних зерен, розташованих рівномірним шаром; б) лусочок; в) тонких паличок, часто вигнутих і на кінці закручених, наприклад, на стеблах цукрової тростини палички воску досягають довжини 0,1 мм.
Восковий наліт знижує інтенсивність транспірації у листя: в дослідах листя евкаліпта, з яких обережно видаляли віск, випарювали води на 30% більше в порівнянні з контрольними.
Восковий покрив може мати і інше значення, особливо для рослин теплих дощових районів. Він робить поверхню органів несмачіваемих і з них легко і швидко стікає вода.
У комахоїдних рослин з родуNepenthesквіти мають вигляд урночек або ковпачків. Їх поверхня зсередини покрита дрібними восковими лусочками, легко відокремлюються при натиску лапок комах. Навіть безкрилі оси, здатні повзати по вертикальній поверхні скла, сідаючи на околицю урни, не можуть втриматися і неминуче падають на дно.
Епідерма листя і стебел рослин, що ростуть зануреними у воду, майже не має кутикули і, тим більше, воскового нальоту. Чим суші місцепроживання рослини, тим кутікулярная плівка більш чітко виражена.
Іноді епідерміс складається з декількох шарів клітини. Припускають, що в цьому випадку епідерміс виконує водозапасающими функцію. Оскільки така епідерміс відзначена переважно у тропічних рослин, які ростуть в умовах мінливої забезпеченості водою, таких як фікуси, бегонії.
У клітинах епідерми можуть утворюватися різні продукти життєдіяльності протопласта. Особливо цікаві так звані цістоліти . Вони формуються в гіпертрофічна розрослися клітинах епідерми і являють собою гроздьевідних кристали вуглекислого вапна. Клітини з цістолітамі є ідіобласти.
Інша особливість зовнішніх стінок клітин епідерми окремих рослин ≈ просочування їх мінеральними солями кальцію і кремнію. У осок кремній відкладається навіть в кутикуле. У деяких випадках клітинні оболонки набувають настільки велику міцність, що хвощі, наприклад, в шкірці яких відкладається кремнезем, використовують для полірування.
Наявність безперервного шару кутикули позбавило б рослини можливості будь-якого газообміну із середовищем, що неминуче призвело б його до загибелі. Тому в процесі еволюції виникли специфічні структури ≈ продихи. Через них і здійснюється повідомлення з зовнішнім середовищем. Там, де немає кутикули, наприклад, у підводних рослин, немає і устьиц. Чим товщі кутикула, тим більшим продихи. Через продихи проходить надзвичайно інтенсивна дифузія водяної пари, кисню і вуглекислого газу.
Кожне продихи складається з пари замикаючих клітин і устьичной щілини, яка представляє собою межклетники. Замикають клітини відрізняються від тих, що оточують їх звичайних епідермальних клітин своєю формою і наявністю хлоропластів. Найчастіше замикають клітини мають бобовидную форму. Крім того, замикають клітини зазвичай мають більш дрібні розміри.
Як правило, замикають клітини оточені так званими побічними клітинами продихів, що відрізняються морфологічно від основних клітин епідерми. Побічні клітини функціонально тісно пов'язані з замикаючими і складають разом устьічнимі апарат (Або устьічнимі комплекс). Побічні клітини продихів дуже різноманітні за формою, для них навіть розроблена спеціальна класифікація, а примітивні вищі рослини взагалі не мають побічних клітин.
Розкриття і закривання продихів представляє надзвичайно важливе явище в житті рослин. Повністю механізм роботи устьічного апарату був виявлений зовсім недавно, але вже з часів Швенденер відомо, що основним фактором тут є зміна тургору (осмотичного тиску) всередині замикаючих клітин.
Розкриттю устьиц, крім того, сприяє нерівномірно потовщені оболонки замикаючих клітин. Внутрішні стінки, оздоблюють устьічнимі щілину, більш товсті, ніж зовнішні. Тому при підвищенні тиску в замикаючих клітинах зовнішні стінки згинаються сильніше і устьичная щілину відкривається.
Зміна тургорного тиску в замикаючих клітинах обумовлено зміною в них концентрації іонів калію. Іони калію закачуються в прикінцеві клітини проти градієнта концентрації. На це потрібна велика кількість енергії, тому що замикають клітини містять численні мітохондрії. Вуглеводи, необхідні для активної діяльності мітохондрій, синтезуються хлоропластами.
При високій концентрації калію вода всмоктується в прикінцеві клітини, їх обсяг збільшується і продихи відкривається.
Відтік іонів калію і відповідно води відбувається пасивно.
Резервуаром іонів калію служать побічні клітини.
У русі устьиц особливе значення має також і радіальна орієнтація целюлозних микрофибрилл в оболонках замикаючих клітин. Ці радіальні міцели дозволяють замикаючим клітинам подовжуватися і водночас не дають їм розширюватися.
У більшості випадків продихи в значно більших кількостях розташовані на нижньому боці листових пластинок, ніж на верхній. В цьому випадку продихи не схильні до прямого впливу сонячних променів і менше нагріваються.
Устячка на верхній стороні листа переважають у трав'янистих рослин, Що мешкають на сильно нагріваються кам'янистих схилах.
І, нарешті, у водних рослин, таких, як латаття, водної лілії, у яких листя розташовані на поверхні води, продихи знаходяться тільки на верхній стороні листа.
Таким чином, кількість продихів і їх локалізація в значній мірі залежить від екологічних умов. В середньому на 1 мм 2 поверхні листа налічується 100-300 устьиц.
У величезної більшості вищих рослин клітини епідерми утворюють вирости ≈ трихоми або волоски (грец. тріхос ≈ волосок). До трихоми відносяться найрізноманітніші вирости епідерми. Деякі з них дійсно нагадують формою волоски, інші мають вигляд сосочків, горбків, гачків, лусочок.
трихоми бувають залізисті і криють . У залізистих трихоми накопичуються екськрети, тому їх відносять до видільної системи.
Крім того, розрізняють волоски одноклітинні і багатоклітинні, мертві і живі.
Мертві волоски позбавлені протопласта, порожнини їх заповнені повітрям, внаслідок чого вони здаються білими. Рослина густо вкрите мертвими волосками, має сивий вигляд. Такі волоски краще відображають сонячні промені і цим зменшують нагрівання і випаровування у рослини.
Форма волосків дуже різноманітна і характерна для того чи іншого виду рослини. Волоски бувають головчатиє, зірчасті, крючковідние, лускаті, гіллясті.
Нерідко трихоми захищають рослини від комах. При цьому, чим густіше опушено рослина, тим рідше комахи відвідують його і використовують в якості їжі і для відкладання яєць.
Від трихоми слід відрізняти емергенци (Лат. emergere - видаватися) ≈ структури, в утворенні яких бере участь не тільки епідерміс, але і глибше розташовані тканини. У деяких рослин, малин, троянд, утворюються емергенци, звані шипами. В освіті шипів у шипшини, наприклад, крім епідерми беруть участь 2 нижче лежачих шару. Від справжніх колючок (метаморфозів органів) емергенци відрізняються безладним розташуванням.
перидерма
У більшості багаторічних рослин осьові органи (корінь і стебло) неухильно наростають в товщину, за рахунок діяльності вторинних меристем. Але первинна покривна тканина ≈ епідерміс не володіє меристематической активністю і не може слідувати за потовщенням органів. Під напором утворюються вторинних тканин вона розривається і слущивается. На зміну їй приходить складна вторинна покривна тканина ≈ перидерма.
Початок утворення перідерми кладеться формуванням вторинної латеральної меристеми ≈ коркового камбію або феллогена .
Феллоген працює на дві сторони. Назовні він відкладає шари пробки (або Фелл ), А всередину органу ≈ живу Паренхімні тканину ≈ феллодерми у.
На поперечному зрізі органу феллоген виглядає у вигляді кільця щільно зімкнутих тонкостінних клітин, що мають, як правило, прямокутні обриси.
Пробка (феллема)
Клітини пробки утворюють правильні радіальні ряди над виробляють їх клітинами феллогена.
характерна особливістьклітин феллеми ≈ суберінізація їх оболонок. жироподібна речовина суберин відкладається у вигляді суцільних пластинок, а самі целюлозні оболонки, крім цього, нерідко дерев'яніють. Оболонки пробкових клітин можуть мати коричневий або жовтий колір.
Пори в клітинах пробки розвинені слабко. У цьому немає необхідності, оскільки живе вміст в клітинах пробки зазвичай рано відмирає і порожнини клітин заповнюються повітрям.
У деяких рослин в клітинах пробки є зернисте вміст, наприклад, біле смолоподобное речовина бетулін у берези або стирол в клітинах дуба. Припускають, що ці речовини мають антисептичні властивості.
У багатьох рослин, наприклад, у беріз, щорічно утворюються нові тонкостінні і товстостінні шари пробки, щось на зразок річних кілець. Саме тому береста легко розшаровується.
Потужність феллеми сильно варіює. Надзвичайно потужну пробку мають знамениті пробкові дуби (темно-зелена пробка).
Пробка функціонує в якості покривної (захисної) тканини. Особливо велика роль пробки у надземних органів. Пробка непроникна для води, тому охороняє стовбури і гілки від висихання.
Рітідом
У більшості деревних порід на зміну вторинної покривної тканини перидерма приходить третинна покривна тканина - кірка або рітідом .
Діяльність коркового камбію не може тривати необмежено довго. Зазвичай через певний період часу його активність згасає і тоді в більш глибоких шарах кори закладається новий шар феллогена, що виробляє пробку. Так, в результаті багаторазового закладання нових прошарків перідерми і утворюється третинна покривна тканина - рітідом. Живі клітини, що потрапили між цими прошарками, гинуть. Таким чином, кірка являє собою складний тканинний комплекс, що складається з чергуються шарів пробки і інших відмерлих тканин.
Мертві тканини, що становлять кірку, розтріскуються під напором наростаючих зсередини тканин, тому на відміну від гладкої пробки, кірка характеризується трещиноватостью.
Особливо яскраво межа між гладкою перидермой (берестой) і тріщинуватої кіркою виражена при підставі стовбурів берези.
Груба, нерідко товста шкірка надійно оберігає стовбури дерев від механічних пошкоджень, різкої зміни температур і навіть лісових пожеж.
механічні тканини
вищі рослиниведуть прикріплений спосіб життя, тому здатність протистояти навантаженням має для них особливе значення. Деякі рослини змушені десятки і навіть сотні років чинити опір бурям, злив, граду і снігу.
Будь-яка рослинна клітина(За рідкісним винятком) має зовнішній скелет у вигляді целюлозної оболонки. Тому, вельми примітно, сто в забезпеченні міцності тіла рослини немає різкого розмежування між різними тканинами. У той час як робота фотосинтезу може виконуватися лише спеціалізованими клітинами, забезпеченими хлоропластами, функцію зміцнення рослинного тіла в тій чи іншій мірі виконують всі клітини і тканини, живі і мертві. Ще раз повторю, що в забезпеченні міцності рослини беруть участь всі клітини і тканини.
Однак якщо для невеликих багатоклітинних організмів(Особливо занурених у воду) наявність тонких оболонок у кожної з клітин виявляється цілком достатнім для забезпечення міцності і підтримки форми, то для великих наземних рослин така опорна система явно недостатня і у них виникли спеціалізовані механічні тканини, які навіть після відмирання живого вмісту продовжують виконувати опорну функцію.
Тіло рослини в цілому видається чимось на зразок залізобетонної конструкції, де механічні тканини відповідають арматурі (тяжам дроту і стержнів), а інші тканини відіграють роль наповнювача (бетону). Саме тому Ф.М. Раздорський вважав більш правильним називати механічні тканини ≈ арматурними .
Розрізняють два основних типи механічних тканин: Коленхіма і склеренхіма .
Коленхіма
Коленхіма (грец. кола- клей), яку розглядають як опорну тканину, складається з товстостінних клітин. Вона дуже близька до паренхімі. Клітини обох тканин містять протопластів з усіма органелами. Для того й іншого тканини характерні первинні і нелігніфіцірованние оболонки клітин. Відмінність між цими тканинами полягає головним чином в тому, що у коленхіми оболонки товщі, а клітини прозенхімних витягнуті в довжину і мають скошені кінці.
Крім того, характерна особливість клітин коленхіми полягає в тому, що їх оболонки нерівномірно потовщені , Що надає цій тканини на поперечному зрізі дуже своєрідний вид.
Залежно від характеру потовщення розрізняють три основних типи коленхіми: 1) кутикової ; 2) пластинчатую і 3) пухку .
У клітинах уголковой коленхіми оболонка сильно потовщується в кутах, де стикається кілька клітин.
Пластинчаста колленхіма, менш поширена, характеризується тим, що потовщені частини оболонок розташовані паралельними шарами, а самі клітини на поперечному зрізі мають форму прямокутників.
Порівняно рідко зустрічається пухка колленхіма. Будова цієї тканини ускладнюється тим, що клітини на ранніх стадіях розвитку роз'єднуються в кутах з утворенням межклетники. Оболонки же товщають саме в місцях прилеглих до межклетники.
Коленхіма пристосована насамперед для виконання функції опори зростаючих листя і стебел. Коленхіма з'являється на ранніх етапах розвитку втечі. Її оболонки пластичні і здатні до розтягування. Тому вона не перешкоджає подовженню органу. Якби в цей час виникали жорсткі тканини, нездатні до розтягування, то і подовження органів було б неможливим. Крім того, колленхіма здатна не тільки пасивно розтягуватись, а й активно рости, завдяки збереженню в клітинах живого вмісту.
Одна з особливостей коленхіми полягає в тому, що вона виконує своє призначення тільки в стані тургору . Якщо молоді пагони втрачають воду, тонкі ділянки оболонок складаються "гармошкою" і пагони в'януть, тобто втрачають пружність, обвисають.
Отже, колленхіма жива тканина, що складається з витягнутих клітин з нерівномірно потовщеними стінками, здатна розтягуватися і виконує свої функції лише в стані тургору клітин.
Для коленхіми характерно периферичний становище у втечі. Вона розташовується або безпосередньо під епідермою, або на відстані одного або декількох шарів від неї. У стеблах колленхіма частіше утворює суцільний кільцевої шар. Вона зустрічається іноді і у формі тяжів в ребрах трав'янистих стебел.
склеренхіма
Склеренхіма ≈ це найбільш поширений в рослинному царстві тип механічної тканини.
Як і колленхіма, вона складається з витягнутих прозенхімних загострених на кінцях клітин, але в іншому помітно від неї відрізняється.
Типова склеренхіма складається з рівномірно потовщених , Щільно зімкнутих клітин. Сформувавшись, клітини склеренхіми зазвичай втрачають живе вміст і їх порожнини заповнюються повітрям; клітинні стінки до цього часу в більшості випадків дерев'яніють ; порівняно рідко вони залишаються чисто целюлозними. Таким чином, склеренхіма виконує свою функцію вже після відмирання протопластів.
Матеріал клітинних стінок склеренхіми має високу міцність і пружністю. По міцності на розрив він близький до будівельної стали, а по пружності навіть перевищує її. Склеренхіма значно перевершує сталь в здатності протистояти динамічним (ударним) навантаженням без деформації.
Склеренхіма мається на вегетативних органах майже всіх судинних рослин. Її немає або вона слабо розвинена в занурених у воду органах.
Зазвичай склеренхімние клітини поділяються на дві групи: 1) волокна і 2) склереїди .
Волокна представляють собою довгі вузькі прозенхімних клітини, довжина яких у багато разів перевищує ширину. Зазвичай волокна мають товсті стінки і дуже вузьку порожнину. Міцність стінок підвищується ще й тому, що фібрили целюлози проходять в них гвинтоподібно, а напрямок витків в шарах чергується.
Волокна вторинного походження відзначаються спеціальними термінами. Назовні від камбію в лубе формуються так звані луб'яні волокна, а всередину від камбію в деревині відкладаються деревинні волокна або лібриформ .
Волокна можуть розміщуватися у вигляді суцільного кільця, окремими тяжами і навіть поодинці.
У практиці луб'яні пучки називають технічним волокном. Луб'яні волокна деяких рослин широко використовуються в промисловості. Міцність і сировинне якість технічного волокна залежать від ступеня зчеплення луб'яних клітин між собою і щільності з'єднання їх в пучки, а також від довжини волокна. Текстильні тканини з довгих тонких і звивистих волокон мають велику міцність.
У виробництві особливо цінні ті рідкісні рослини, у яких волокна не дерев'яніють, наприклад, льон.
Назву лише найбільш відомі волокнисті рослини і вироби з них виготовляються: конопля(Cannabis sativa)≈ канати і мотузки; джут(Corchorus capsularis)≈ канати, мотузки і грубі тканини; кенаф(Hibiscus cannabinus)≈ грубі тканини; льон(Linum usitatissimum)≈ тканини; рами (Bochmeria nivea)≈ тканини.
склереїди
Склереїди не мають форму волокон і сильно варіюють за формою. Зазвичай за формою клітин їх і класифікують.
Найбільш поширеними є ізодіаметріческіе округлі склереїди, звані брахісклереідамі або кам'янистими клітинами.
Брахісклереіди зустрічаються в шкаралупі плодів ліщини, жолудя; в кісточках плодів сливових, волоського горіха; в м'якоті плодів груші, айви; в шкірці насіння кедрової сосни.
Брахісклереіди зустрічаються і в підземних органах ≈ в корі кореневищ півоній, в коренях хрону, бульбах жоржини.
Остеосклереіди мають длінноціліндріческую форму з розширеннями на кінцях, що нагадують сочленовой головки кісток.
Астросклереіди або зірчасті склереїди мають гіллясту форму. Найчастіше астросклереіди зустрічаються в листі шкірястої консистенції. Вони надають листовим пластинках додаткову міцність.
Склереїди можуть утворювати суцільні групи, тканинну масу, як в шкаралупі плодів. Можуть вони зустрічатися і поодинці, у вигляді ідіобласти, як, наприклад, в листі.
Організація тіла рослини з точки зору будівельно-механічних принципів
Прикріплені до місця рослини не мають можливості ховатися від дії стихій. При цьому рослини нерідко мають дуже великими розмірамиі майже завжди ≈ величезною зовнішньою поверхнею. Вони піддаються енергійному впливу різноманітних механічних навантажень. До того ж багато навантаження існують десятки і сотні років, тобто є багаторазовими.
Тому рослини мають вражаючу здатність протистояти різного роду механічних навантажень. Тонка соломина підтримує важкий колос і листя, розгойдується при поривах вітру і не ламається.
Ця житнє соломина борг час викликала здивування ботаніків. Її порівнювали то з Ейфелевою вежею, то з високими димарями. У соломини висота в 500 разів перевищує діаметр при основі, а інженерні споруди за цим показником помітно програють.
Доцільність будови рослин з точки зору механіки намагався пояснити ще Галілей. Його цікавило питання, як змінюються пропорції тіла організмів при значній зміні їх розмірів. Багато уваги цим питанням приділяли Грю і Гук. Однак лише два століття по тому в 1874г німецький ботанік Швенденер детально розглянув розподіл механічних тканин в тілі рослини з точки зору інженерної теорії опору матеріалів.
Було висунуто положення, яке стверджує, що органи рослин будуються відповідно до принципів досягнення міцності при економною витраті матеріалу.
Розглянемо це положення більш детально.
Якщо стрижень, який відчувають на міцність, покласти на дві опори і навантажити, то він прогнеться. При цьому його нижня сторона буде розтягуватися, тобто протидіяти розриву. Навпаки, верхня сторона буде протидіяти раздавливанию і стискатися. Матеріал же знаходиться в центрі стержня залишиться в цьому відношенні нейтральним.
Таким чином, з точки зору інженерних розрахунків матеріал доцільно зосередити у верхній і нижній частинах стрижнів, де він буде виконувати найбільше навантаження. У центрі ж з метою економії вжити лише в тій мірі, щоб запобігти стисненню конструкції в поперечному напрямку.
Відповідно до цих принципів інженери встановили найбільш економічну і доцільну конструкцію у вигляді двотаврової балки, яка застосовується для перекриттів.
Швенденер показав, що в листі рослин механічні тканини по розташуванню дуже нагадують двотаврові балки.
Стебло піддається вигинів в різних напрямкахі його можна порівняти з вертикальної трубою. З метою найбільшої економії і міцності механічні елементи повинні бути віднесені до периферії такої конструкції. Дійсно, в стеблах колленхіма і склеренхіма найчастіше розташовуються безпосередньо під епідермою або близько до поверхні. Центр стебла зазвичай зайнятий тонкостінної паренхіми або навіть має велику порожнину. Соломіна злаків є типовою порожнистої трубою.
Корню, оточеному грунтом, не загрожує небезпека вигину і зламу. Корінь виконує іншу механічну задачу, він "заякорюють" рослини в грунті і протидіє напруженням, які прагнуть висмикнути його, тобто протидіє розриву. У відповідність з цим, механічні тканини розміщуються в центрі кореня.
Істотний внесок в проблему, яка розглядається вніс вітчизняні ботанік В.Ф.Раздорскій. Перш за все Раздорський показав, що розглянуті нами принципи розподілу механічних тканин в різних органах рослини реалізуються не настільки однозначно. Справа в тому, що рослини впадають впливу механічних навантажень 2-х категорій, що пред'являють до них абсолютно протилежні вимоги, навантажень: 1) статичним і 2) динамічним .
До перших належить вплив сили тяжіння, власної ваги. Іноді статичні навантаження помітно збільшуються, наприклад, при осіданні снігу в гілках крон.
До механічних факторів динамічного (ударного) роду можна віднести пориви вітру, удари крапель дощу і граду.
Схема, запропонована Швенедером, в більшій мірі відповідає протидії статичних навантажень.
Однак, з точки зору динаміки, як показав Раздорський, органи рослини повинні працювати подібно пружинам вигину, здатним щоразу повертатися в початковий стан після зняття навантаження. Тому стовбур дерева, розгойдується під вагою крони, має конструкція не порожнистої жорсткої труби, а суцільний пружною пружини.
Крім того, "інженерні вимоги" рослин помітно змінюються в ході онтогенезу. У молодого проростка чітко проявляється периферична тенденція розвитку механічних тканин. У цій стадії рослина тягнеться до світла і його завдання якомога швидше досягти максимальної висоти з можливою економією матеріалу.
Але, вибравшись до світла, рослина починає гілки і піддається більш сильному впливу вітру і інших динамічних факторів. Відповідно змінюється і розподіл механічних тканин: більшою мірою зміцнюється центр.
Раздорський же вперше звернув увагу на схожість рослин з так званими комплексними спорудами, до розряду яких головним чином відносяться залізобетонні конструкції.
В органах рослини бетону відповідає основна маса м'яких і тонкостінних тканин, а каркасу (арматурі) ≈ механічні тяжі.
Але рослинні конструкції стоять на вищому щаблі, ніж технічні споруди. Бетон руйнується вже при незначному подовженні в 0,01%. Тому в залізобетоні високі механічні якості заліза використовуються далеко не повністю. У рослинних конструкціях основна маса здатна сильно деформуватися, тканини заповнення можуть зруйнуватися лише після розриву тяжів арматури. Тому в органах рослин опір арматури використовується повністю.
Крім того, залізо має властивість плинності, тобто залізний прут або витягується, або пучітся при стисненні, без повернення в початковий стан. Рослинна арматура плинністю не володіє.
Однак природа завжди різноманітніше, ніж наші судження про неї. Деякі рослини будуються як би наперекір інженерним принципам. Наприклад, південно-африканські Клейн(Kleinia- з сімейства складноцвітих) мають стебла тонкі біля основи і товщають на вершині. В результаті гілки відламуються при найменшому вітрі. Відламалися пагони служать для вегетативного розмноження.
[Попередня лекція] [Зміст] [Наступна лекція]КЛАСИФІКАЦІЯ ТКАНИН
Царство рослин складається з одноклітинних і багатоклітинних організмів.
Якщо це одноклітинний організм, то в його єдиній клітині відбуваються всі процеси життєдіяльності; в багатоклітинних організмі різні клітини об'єднуються в групи однаково функціонуючих клітин - тканини.
тканини- стійкі, закономірно повторювані комплекси клітин, подібні за походженням, будовою і пристосовані до виконання однієї або кількох функцій.
Залежно від основної функції розрізняють кілька груп рослинних тканин.
1. Освітні тканини,або меристеми,- мають здатність до поділу і формування всіх інших тканин.
2. Покривні тканини:
первинні;
вторинні;
Третинні.
3. Основні тканини- складають більшу частину тіла рослини. Розрізняють такі основні тканини:
Асиміляційні (хлорофіллоноснимі);
запасають;
Повітроносні (аеренхіма);
Водоносні.
4. механічні тканини(Опорні, скелетні):
Коленхіма;
Склеренхіма.
5. Провідні тканини:
Ксилема (деревина) - тканина висхідного струму;
Флоема (луб) - тканина спадного струму.
6. Видільні тканини:
зовнішні:
Залізисті волоски;
Гідатоди - водяні продихи;
нектарники;
внутрішні:
Видільні клітини з ефірними маслами, смолами, дубильними речовинами;
Багатоклітинні вмістилища виділень, молочні судини.
За здатністю клітин до поділу розрізняють два типи тканин: освітні,або меристеми,і постійні- покривні, видільні, основні, механічні, провідні.
тканина називають простий,якщо всі її клітини однакові за формою і функцій (паренхіма, склеренхіма, колленхіма). складнітканини складаються з клітин, неоднакових за формою, будовою і функціями, але пов'язаних загальним походженням (наприклад, ксилема, флоема).
Існує також класифікація тканин, заснована на їх походження (онтогенетическая). Відповідно до цієї класифікації розрізняють первинні і вторинні тканини. З первинної меристеми, що знаходиться на верхівці втечі і кінчику кореня, а також з зародка насінини формуються первинні постійнітканини (епідерміс, колленхіма, склеренхіма, ассимиляционная тканину, епіблема). Клітини постійних тканин нездатні до подальшого поділу. З клітин спеціалізованої меристеми - прокамбію - формуються первинні провіднітканини (первинна ксилема, первинна флоема).
З вторинної меристеми - камбію - формуються вториннітканини: вторинна ксилема, вторинна флоема; з феллогена утворюються пробка, феллодерми, чечевички, що виникають при потовщенні стебла і кореня. Вторинні тканини, як правило, зустрічаються у голонасінних і дводольних покритонасінних рослин. Потужний розвиток вторинних тканин - деревини і лубу характерно для деревних рослин.
ОСВІТНІ ТКАНИНИ
Ріст рослин на відміну від зростання тварин має дві особливості. По-перше, рослина має унікальну властивість необмеженого зростання: воно росте протягом усього життя, утворюючи нові клітини. По-друге, рослина характеризується локалізованість зростання в певних місцях - точках зростання. У точках зростання перебувають освітні тканини - меристеми, основною функцією яких є створення нових клітин шляхом поділу. Крім того, у рослин можливо новоутворення меристем. У таких випадках деякі постійні тканини проходять дедіфференціровка і набувають здатність ділитися. На цьому, наприклад, засновані регенерація і вегетативне розмноження рослин.
освітні тканинизавдяки постійному мітотичного поділу їх клітин забезпечують освіту всіх тканин рослини, тобто фактично формують його тіло. Будь-яка клітина в своєму розвитку проходить три стадії: ембріональну, зростання і стадію диференціації (тобто придбання клітиною певної функції). У міру диференціації зародка первинна меристема зберігається тільки на верхівці майбутнього втечі (в конусі наростання) і на кінчику кореня - апікальні (верхівкові) меристеми.Зародок будь-якої рослини складається з клітин меристеми. Клітини первинної меристеми мають густу цитоплазму і великі ядра, інтенсивно діляться митозом. У гіалоплазме багато дифузно розкиданих рибосом, пропластид, мітохондрій і диктиосом. Вакуолей трохи, і вони дрібні. Провідні тканини утворюються з меристеми, що має прозенхімних форму і великі вакуолі, - прокамбію і камбію. Клітини прокамбію в поперечному перерізі багатокутні, клітини камбію - прямокутні.
Клітини, що зберігають свої меристематические властивості, продовжують ділитися, утворюючи все нові і нові клітини, звані ініціалами.Частина дочірніх клітин диференціюється, перетворюючись в клітини різних тканин, їх називають похідними ініціали.Клітини ініціали можуть ділитися невизначено багато разів, а похідні ініціали діляться один або кілька разів і перетворюються в постійні тканини.
За походженням розрізняють первинні і вторинні меристеми (табл. 2.1).
первинні меристеми
Первинні меристеми відбуваються безпосередньо з меристеми зародка і мають здатність до поділу. Відповідно до положення в рослині первинні меристеми можуть бути верхівковими (апікальними) (рис. 2.2), вставними (інтеркалярний) і бічними (латеральний).
Верхівкові (апікальні) меристеми- такі меристеми, які розташовуються у дорослих рослин на верхівках стебел і кінчиках коренів і забезпечують зростання тіла в довжину. У стебел в конусі наростання виділяють два меристематических шару: туніку, з якої утворюються покривна тканина і периферична частина первинної кори, і корпус, з якого утворюються внутрішня частина первинної кори і центральний осьовий циліндр (рис. 2.3).
У кінчику кореня розрізняють три шари:
1) дерматоген, з якого утворюється первинна покровно-усмоктувальна тканина - ризодерма;
2) періблему, з якої розвиваються тканини первинної кори;
3) плером, який утворює тканини центрального осьового циліндра (рис. 2.4).
Мал. 2.2. Верхівкова меристема втечі: 1 - конус рослини; 2 - зачатки листя; 3 - зачаток бічній нирки
Бічні (латеральні) меристемиза походженням можуть бути первинними і вторинними, на поперечному зрізі осьових органів мають вигляд кілець. Прикладом первинної бічній меристеми служать прокамбій і перицикл. з прокамбіюформуються камбій і первинні елементи судинно-волокнистих пучків (первинна флоема і первинна ксилема), при цьому клітини прокамбію безпосередньо диференціюються в клітини первинних провідних тканин.
Бічні меристеми розташовуються паралельно поверхні органу і забезпечують зростання осьових органів в товщину.
Вставні (інтеркалярние) меристемичастіше первинні і зберігаються у вигляді окремих ділянок в зонах активного росту в різних частинах рослини (наприклад, в підставі черешків листя, у підстав междоузлий). У підставі междоузлий у злаків діяльність цієї меристеми веде до подовження міжвузлів, що забезпечує ріст стебла в довжину.
вторинні меристеми
До вторинних відносять бічні і ранові меристеми.
Бічні (латеральні) меристемипредставлені камбіємі феллогеном.Вони формуються з промерістем (прокамбію) або постійних тканин шляхом їх дедіфференціровкі. Клітини камбію діляться перегородками, паралельними поверхні органу (периклінальних). З клітин, відкладених камбієм назовні, розвиваються елементи вторинної флоеми, а з клітин, відкладених всередину, - вторинної ксилеми. Камбій, що виник з постійних тканин шляхом дедіфференціровкі, називають добавочньм.За будовою і функції він не відрізняється від камбію, що виник з промерістем. Феллоген формується з постійних тканин, розташованих в субепідермальной шарах (під епідермою). Ділячись периклінальних, феллоген відокремлює назовні майбутні клітини пробки (феллеми), а всередину - клітини феллодерми. Таким чином, феллоген формує вторинну покривну тканину - перидерму. Бічні меристеми розташовуються паралельно поверхні органу і забезпечують зростання осьових органів в товщину.
ранові меристемиутворюються при пошкодженні тканин і органів. Навколо пошкодження живі клітини дедіфференціруются, починають ділитися і тим самим перетворюються у вторинну меристему. Їх завдання - утворити щільну захисну тканину, що складається з паренхімних клітин, - каллюс.Ця тканина білуватого або жовтуватого кольору, її клітини мають великі ядра і досить товсті клітинні стінки. Каллюс виникає при щепленнях, забезпечуючи зрощення прищепи з підщепою, і в підставі черешків. У ньому можуть закладатися додаткове коріння і нирки, тому його використовують для отримання культури ізольованих тканин.
покривних тканин
На поверхні всіх органів рослин перебувають покривні тканини. Основна їхня функція - захисна. Вони захищають внутрішні тканини від несприятливих факторів: механічних впливів, різкої зміни температур, проникнення шкідників і патогенних мікроорганізмів, зайвого випаровування і осушення. Крім захисної, покривні тканини виконують і інші функції - газообміну, транспірації, виділення, поглинання та ін.
Розрізняють первинну, вторинну і третинну покривні тканини.
Первинна покривна тканина
Вона покриває листя, квітки, плоди, а також стебла при їх первинному будову.
Епідерма виконує в основному функції покривних тканин. З поліфункціональність пов'язано різноманітність її структур, т. Е. Шкірка - складна тканина. Вона складається з наступних компонентів:
1. Основна тканина, клітини якої утворюють велику частину її поверхні.
2. Устячка і пов'язані з ними побічні клітини.
3. Різні вирости (волоски, трихоми).
До первинної покривноїтканини відносять епідерму власне епідермальних, околоустьічних, замикаючих клітин продихи і трихом.
Клітини основної тканини шкірки паренхімні, щільно зімкнуті, іноді мають звивисті стінки, що сприяє більш міцному їх з'єднанню. Епідерма органів, витягнутих у довжину (стебла, черешки, ланцетоподібні листя), складається з клітин, подовжених паралельно поздовжньої осі органу. Бічні і внутрішня стінки клітин тонкі целюлозні.
Пектинові речовини і целюлоза, що входять до клітинну стінку, Можуть піддаватися ослизнення з утворенням слизейі камеді.Вони являють собою полімерні вуглеводи, родинні пектиновим речовин, і відрізняються здатністю до сильного набухання при зіткненні з водою. Камеді в набряклому стані клейкі і можуть витягуватися в нитки, тоді як слизу сильно розпливаються і в нитки не витягуються. Пектинові слизу зустрічаються у представників сімейств лілейних, хрестоцвітих, мальв, липових, розоцвітих, на відміну від целюлозних слизу, що зустрічаються набагато рідше (наприклад, у орхідних).
Газообмін між внутрішніми тканинами рослин і зовнішнім середовищем, а також випаровування води в епідермі здійснюються через спеціальні освіти - продихи.
устячка являють собою високоспеціалізовані освіти епідерми, що складаються з двох замикаючих клітин бобовидной форми і устьичной щілини (своєрідного межклетники між ними). Є головним чином в листі, але зустрічаються і в стеблі (рис. 2.6).
Устячка розташовані на нижньому боці аркуша, але у водних рослин з плаваючим листям вони знаходяться тільки на верхній стороні листа. За формою клітин епідерми листа і розташуванню устьиц можна відрізнити однодольное рослина від двудольного (рис. 2.8). Власне епідермальні клітини листа дводольних рослин в обрисах хвилясті (рис. 2.9), а в однодольних вони витягнуті, ромбічної форми.
Щілина може розширюватися і звужуватися, регулюючи транспирацию і газообмін. Під щілиною розташована велика дихальна порожнина (межклетники), оточена клітинами мезофіла листа.
Замикають клітини оточені околоустьічнимі клітинами, які спільно утворюють устьічнимі комплекс(Рис. 2.7). Розрізняють такі основні типи устьічних комплексів:
1) аномоцітний(Безладний) - замикають клітини не мають яскраво виражених околоустьічних клітин; характерний для всіх вищих рослин, за винятком хвойні;
2) анізоцітний(Неравноклеточний) - замикають клітини продихи оточені трьома околоустьічнимі клітинами, одна з яких набагато більше (або менше) інших;
3) парацітний(Паралельно-клітинний) - одна околоустьічная клітина (або більше) розташована паралельно замикаючим;
4) діацітний(Перехресно-клітинний) - дві околоустьічние клітини розташовані перпендикулярно замикаючим;
5) тетрацітний(Від грец. tetra- чотири) - головним чином у однодольних;
Мал. 2.7.Основні типи устьічного апарату: 1 - аномоцітний (у всіх вищих рослин, крім хвощів); 2 - діацітний (у папоротей і квіткових); 3 - парацітний (у папоротей, хвощів, квіткових і гнетових); 4 - анізоцітний (тільки у квіткових); 5 - тетрацітний (головним чином у однодольних); 6 - анціклоцітний (у папоротей, голонасінних і квіткових)
Механізм роботи продихів обумовлений осмотичним властивостями клітин. При висвітленні поверхні листа сонцем в хлоропластах замикаючих клітин відбувається активний процес фотосинтезу. Насичення клітин продуктами фотосинтезу, цукрами тягне за собою активне надходження в клітини іонів калію, внаслідок чого концентрація клітинного соку в замикаючих клітинах зростає. Виникає різниця концентрації клітинного соку околоустьічних і замикаючих клітин. В силу осмотических властивостей клітин вода з околоустьічних клітин надходить в прикінцеві, що веде до збільшення обсягу останніх і різкого зростання тургору. Потовщення «черевних» стінок замикаючих клітин, звернених до устьичной щілини, забезпечує нерівномірний розтяг клітинної стінки; замикають клітини набувають виражену бобовидную форму, і устьичная щілину відкривається. При зниженні інтенсивності фотосинтезу (наприклад, ввечері) зменшується утворення цукрів в замикаючих клітинах. Приплив іонів калію припиняється. Концентрація клітинного соку в замикаючих клітинах знижується в порівнянні з околоустьічнимі. Вода шляхом осмосу йде з замикаючих клітин, знижуючи їх тургор, в результаті вночі устьичная щілину закривається.
Клітини епідерми щільно зімкнуті між собою, завдяки цьому епідерміс виконує цілий ряд функцій:
Перешкоджає проникненню хвороботворних організмів всередину рослини;
Захищає внутрішні тканини від механічних пошкоджень;
Регулює газообмін і транспірацію;
Через неї виділяються вода, солі;
Може функціонувати як усмоктувальна тканина;
бере участь в синтезі різних речовин, сприйнятті подразнень і в русі листя.
трихоми - різні за формою, будовою і функціями вирости клітин епідерми: волоски, лусочки, щетинки і т.п. Їх поділяють на криють і залізисті. Залізисті трихоми,на відміну від криють, мають клітини, що виділяють секрет. Кроющие волоски,утворюючи на рослині шерстистий, повстяний чи іншої покрив, відображають частину сонячних променів і тим самим зменшують транспірацію. Іноді волоски знаходяться тільки там, де розташовані продихи, наприклад, на нижньому боці листків мати-й-мачухи. У деяких рослин живі волоски збільшують загальну випаровує поверхню, що сприяє прискоренню транспірації.
Розміри трихом значно варіюють. Найбільш довгі трихоми (до 5-6 см) покривають насіння бавовнику. Кроющие трихоми мають форму простих одне або багатоклітинних, розгалужених або зірчастих волосків. Кроющие трихоми можуть тривалий час залишатися живими або швидко відмирати, заповнюючи повітрям (рис. 2.10).
Від трихом, що виникають тільки за участю епідермальних клітин, відрізняються емергенци, в утворенні яких беруть участь і більш глибоко розташовані тканини субепідермальной шарів (рис. 2.11).
Вторинна покривна тканина
У міру зростання і розвитку рослин епідерміс змінюється вторинної покривної тканиною - перидермой. Зазвичай в кінці літа клітини шкірки стебла, або живі клітини, що лежать під шкіркою, набувають здатність ділитися і перетворюються на вторинну освітню тканину- корковий камбій, або феллоген. Клітини феллогена діляться паралельно поверхні і відкладають назовні численні шари пробки (феллеми), а всередину - один-два шари живої тканини феллодерми. Таким чином, виникає комплекс тканин - пробка, феллоген, феллодерми, який називають перидермой.
власне покривну функціюв цьому комплексі виконує пробка, а корковий камбій, що складається з одного шару клітин, постійно продукує її. Пробка складається з мертвих клітин, у яких клітинна стінка просякнута жироподібним речовиною суберином. Клітини розташовані рівними рядами, мають прямокутну форму (на поперечному зрізі), щільно прилягають один до одного, формуючи багатошаровий футляр. Пробка охороняє внутрішні живі тканини від втрати вологи, різких температурних коливань і проникнення мікроорганізмів.
Мал. 2.12.Будова перідерми: 1 - залишки епідерми; 2 - пробка (феллема); 3 - феллоген; 4 - феллодерми
Живі тканини, що лежать під пробкою, потребують газообмене і видаленні надлишку вологи. Саме тому під Устьіце внаслідок поділу субепідермальной шарів (ще до появи перідерми), а в подальшому і феллогена відкладаються живі, пухко розташовані, з безліччю межклетников паренхімні клітини, звані виконує тканью, яка розриває епідерму і створює можливість газообміну і транспірації з зовнішнім середовищем. Це структурне утворення називають чечевички(Рис. 2.12).
Чечевички, мають вигляд невеликих горбків, чітко виділяються на поверхні пагонів дерев і чагарників (рис. 2.13).
Мал. 2.13.Перидерма і чечевички (стебло бузини): 1 - залишок епідерми; 2 - виконує тканину чечевички; 3 - перидерма: 3а - пробка; 3б - феллоген; 3в - феллодерми
На стовбурах берези їх залишки спостерігаються у вигляді характерних поперечних чорних смужок, у осики вони приймають форму ромбів.
Третинна покривна тканина
У більшості дерев з часом перидерма замінюється - кіркою.
Кірка (рітідом)є третинну покривну тканину, яка утворюється у багаторічних рослин в корені, стеблі, кореневище. Щороку в більш глибоких шарах закладається новий шар феллогена і утворюється перидерма. Зовнішній шар перідерми - пробка - ізолює всі вищележачі тканини, в результаті чого вони відмирають. Таким чином, сукупність численних перидерм з відмерлими між ними тканинами і є кірка (рис. 2.14).
Мал. 2.14.Кірка дуба: 1 - шари пробки; 2 - волокна; 3 - залишки первинної кори; 4 - друзи оксалату кальцію
Перідерми можуть закладатися концентричними колами. При цьому зовнішні шари кірки під напором зсередини. тріскаються і слущиваются у вигляді поздовжніх смуг. Така кірка називається кольчатой (виноград, кипарис, евкаліпт). Однак частіше нові перідерми закладаються не суцільним кільцем, а увігнутими дугами. При цьому зовнішні шари кірки слущиваются у вигляді лусок. Це луската кірка (дуб, в'яз, береза, платан).
Кірка служить більш надійним захистом для рослин, ніж перидерма, оберігаючи їх від перегріву і опіків у разі лісових пожеж.
ОСНОВНІ ТКАНИНИ
Основні тканини (паренхіми)складають більшу частину всіх органів рослин. Вони заповнюють проміжки між провідними і механічними тканинами і присутні у всіх вегетативних та генеративних органах. Ці тканини утворюються за рахунок диференціювання апікальних меристем і складаються з живих паренхіматозних клітин, різноманітних за будовою і функціями. Розрізняють асиміляційну, запасаючу, воздухоносную і водоносну паренхіми.
В асиміляційної, або хлорофіллоносной, паренхімі здійснюється фотосинтез.
Паренхіма представлена живими паренхімними тонкостінними клітинами, в протопласті яких присутні хлоропласти. Виділяють три типи асиміляційні тканин: столбчатая, губчаста і складчаста. Всі вони містяться, як правило, в пластинках листків.
Стовпчаста, або палісадна, - основна фотосинтезуюча тканину в рослині. Клітини її мають циліндричну форму, щільно зімкнуті і розташовуються в листі перпендикулярно верхній епідермі. Зазвичай вони утворюють один шар, рідше два-три. стовпчасті клітинимістять велику кількість хлоропластів, а їх декілька витягнута формасприяє відтоку продуктів фотосинтезу.
Губчаста, або пухка, тканинутакож знаходиться в листі, зазвичай під столбчатой. Вона містить багато межклетников, про що свідчить її назва. Клітини мають округлу або лопатеву форму. Хлоропластів в них менше, ніж в клітинах столбчатой тканини. Важливими функціями губчастої хлоренхіми, поряд з фотосинтезом, є газообмін і транспірація.
складчаста паренхімазустрічається в основному в хвої і листі деяких злаків. Оболонка її клітин утворює всередину складки, які збільшують поверхню оболонки, а значить, і пристенного шару цитоплазми, що містить хлоропласти. Іноді складки утворюються тільки на стінці, зверненої до поверхні органа (бамбук, анемона).
Усмоктувальні, що поглинають. Воду і поживні речовини, необхідні для своєї життєдіяльності, рослини поглинають з навколишнього середовища. Водорості та вищі водні рослини всмоктують їх всією поверхнею свого тіла. У вищих рослин, що живуть на суші, є для цього спеціалізовані поглинають, або всмоктувальні, тканини, які ще називають абсорбційними. До них відносять Ризоїди і поглинають воду волоски мохів, волосконосний шар кореня, спеціальні поглинають воду волоски в епідермі рослин, які ростуть в жаркому і сухому кліматі, покривну тканину повітряних коренів, а також щиток - видозмінену сім'ядолю однодольних.
У мохів відсутні коріння, вони поглинають воду в основному за допомогою ризоидов, що представляють собою вирости шкірки, які відділені від її клітин перегородкою. Часто Ризоїди складаються з однієї тонкостінної подовженою клітини із закругленою верхівкою. Коли верхівка стикається з субстратом, вона утворює вирости - розгалуження. Сфагнові мохи містять спеціальні поглинають воду гіалінові клітини. На стеблах вони утворюють покрив з декількох шарів, а в листочках розташовуються між вузькими хлорофіллоноснимі клітинами. Гіалінові клітини великі, мертві, їх стінки мають спіральні потовщення і дрібні отвори, що відкриваються назовні. Через отвори капілярним шляхом вода надходить в гіалінові клітини, заповнюючи їх. Саме цим пояснюється велика вологоємність мохів: вони утримують кількість води, у багато разів перевищує їх масу.
Волосконосний шар є покривною тканиною кореня в зоні всмоктування вище точки зростання. Він носить назву ризодерма, або епіблеми. Клітини цього шару утворюють вирости - кореневі волоски. Вони живі, з тонкими целюлозними оболонками, пристінним шаром цитоплазми і великий центральної вакуолью. Ядро зазвичай розташоване в тій частині цитоплазми, яка знаходиться в волосині, туди ж проникає і вакуоль.
У одних рослин кожна клітина ризодерма утворює волоски, у інших - в ній є клітини двох типів - тріхобласти, Що утворюють волоски, і атріхобласти, Їх не мають. Причому в коренях різних рослин волоски формуються неоднаково.
До поглинає відноситься веламен- покривна тканина повітряних коренів рослин-епіфітів, які використовують в якості опори стовбури і гілки дерев. Коріння цих рослин перебувають не в грунті, а в повітрі. Веламен складається з декількох шарів мертвих паренхімних клітин, оболонки яких мають спіральні або сітчасті потовщення. В оболонці знаходяться наскрізні отвори, через які в клітини Велам капілярним шляхом проникає вода (дощ, роса)
запасающая паренхіма переважає в стеблі, корені, кореневища. У клітинах цієї тканини відкладаються запасають речовини: білки, жири, вуглеводи.
Рослини відкладають в запас велику кількість органічних речовин. Накопичення і зберігання поживних речовин відбувається в запасающих тканинах. Клітини цих тканин зазвичай тонкостінні або рідше товстостінні, щільно зімкнуті або з межклетниками. Їх оболонки мають прості округлі пори. Іноді запасають клітини дуже великі, особливо в соковитих плодах. Їх можна бачити неозброєним оком, наприклад, на розломі яблука або кавуна.
Запас поживних речовин може знаходитися в клітці в протопласті, в вакуолі, рідше в клітинній оболонці. У протопласті - цитоплазмі, пластидах, сферосомах - речовини можуть відкладатися в твердому (крохмаль, білок), рідкому (масла) і розчиненому стані. Так, в цитоплазмі зазвичай накопичуються білки у вигляді кристалів і алейронових зерен (насіння квасолі, сої, гороху, рицини), крапельок масла (насіння олійних рослин, плоди маслини), в пластидах - крохмаль у вигляді крохмальних зерен (насіння злаків, бобових, бульби картоплі), рідше білки і масла, в сферосомах - масла. У вакуолях в розчині містяться цукри (коренеплоди цукрових буряків, м'якоть соковитих плодів), розчинні полісахариди, наприклад інулін (коріння і бульби топінамбура, жоржини, цикорію, кульбаби). У клітинній оболонці запасні речовини представлені геміцелюлозами, що використовуються в насінні деяких рослин при проростанні (люпин, пальми).
У однорічних рослин запасають тканини знаходяться в основному в насінні і плодах. Вони витрачаються при проростанні і зростанні проростка до його позеленіння і переходу до фотосинтезу. У багаторічних рослин запасні речовини відкладаються не тільки в насінні і плодах, а й в вегетативних органах - корі, деревині і серцевині стебла, в корі і деревині кореня. Ці речовини використовуються рослиною при розпусканні бруньок навесні, зростанні молодих пагонів і коренів. Крім того, у багатьох рослин є спеціалізовані запасають органи - кореневища, бульби, цибулини.
воздухоносной паренхіма, або аеренхіма, складається з живих клітин і великих повітроносних порожнин (межклетников), що представляють собою резервуари для запасу газоподібних речовин. Ці порожнини оточені клітинами основної паренхіми (хлорофіллоносной або запасающей). Аеренхіма добре розвинена у водних рослин в різних органах і може зустрічатися у сухопутних видів. Головне призначення аеренхіма - участь в газообміні, а також забезпечення плавучості рослин (рис. 2.15).
Під покривною тканиною слід розуміти комплекс клітин, розташованих зовні всіх органів рослини.
При спробі дати покривної тканини більш точне визначення, відповідне характеру кожного окремого випадку, (зустрічається ряд труднощів. Покривні тканини різних органів дуже різноманітні за будовою і виконуваних функцій. Покривні тканини надземних органів - листа, стебла - служать для захисту від висихання і пошкодження внутрішніх, більше ніжних тканин, виконують функцію виділення, за їх посередництвом здійснюється газообмін з навколишнім середовищем та ін. Покрови же коренів, особливо закінчень їх, поглинають воду з розчиненими в ній мінеральними речовинами, сприяють розчиненню деяких твердих частинок грунту і захищають внутрішні тканини кореня від механічних пошкоджень . Покрови осьових органів у багаторічних рослин до кінця першого вегетаційного періоду в більшості випадків починають сильно змінюватися: покриви першого року вегетації відмирають, а з них або з інших тканин, розташованих в безпосередній близькості, утворюється покривна тканина іншого типу як за структурою, так і за ряд у додаткових функцій. Отже, необхідно розрізняти первинні покривні тканиниі вторинні покривні тканини. Останні або виникають знову, або представляють собою видозміну структури первинної тканини.
Первинна покривна тканина всіх рослинних органів називається шкіркою, або епідермісом.
Епідерміс утворюється з поверхневого шару первинної меристеми, званого протодерми. Епідерміс складається з клітин, як правило, ізодіаметріческого характеру і завжди з живим вмістом. При відмирання протопласта епідермальних клітин останні перестають служити покровом і захистом перебувають за ними тканин і замінюються інший покривною тканиною.
Якщо розглядати епідермальні клітини зовні, то можна переконатися, що обриси їх в різних органах і у різних рослиндуже різні. Обриси епідермальних клітин формуються у великій залежності від співвідношення в швидкості росту органу в цілому і його поверхні. Наприклад, клітини епідермісу листка з широкою пластинкою, однаково розростається як в довжину, так і в ширину, мають звивисті обриси; а клітини епідермісу стебла молодого рослини, сильно витягуватися в довжину, значно більш витягнуті і мають менше звивисті стінки, ніж клітини епідермісу листка того ж рослини. Лінійні листя однодольних рослин відрізняються довгими епідермальними клітинами.
Клітини епідермісу дуже міцно з'єднані один з одним, значно міцніше, ніж з клітинами підстильної епідерміс тканини. Внаслідок цього епідерміс часто легко відривається від інших тканин, але сам не розривається і здирається відносно великими шматками. Епідерміс зазвичай не має межклетников, але бувають і виключення. Наприклад, межклетники зустрічаються в епідермісі пелюсток. Крім того, в епідермісі інших надземних, переважно зелених, органів є щілиноподібні межклетники, оточені спеціальними замикаючими їх клітинами, - продихи.
У молодих епідермальних клітинах майже вся порожнину клітини заповнена протопластом з чітко видним округлим клітинним ядром і досить помітними, хоча і дрібними, лейкопластах. У міру розростання клітини і збільшення розмірів вакуолей цитоплазма займає постінному положення, і ядро, якщо воно розташоване не в центрі клітини, а в цитоплазмі, притиснутою до оболонки, сплющується і витягується.
У цілком виросла епідермальній клітці вакуоля займає майже всю клітину, вона наповнена клітинним соком, Безбарвним або забарвленим розчиненими в ньому пігментами. Протопласт у вигляді дуже тонкого шару, нерідко з працею помітного, притиснутий до оболонки. Часто в такій клітці пластиди руйнуються, абсолютно зникаючи. Це спостерігається переважно у світлолюбних трав'янистих рослин. Втім і у таких рослин іноді в епідермальних клітинах нижнього боку листа є хлорофілові зерна. У рослин, які ростуть в тіні, хлоропласти часто є у всіх епідермальних клітинах. У деяких рослин пластиди в епідермісі листя зберігаються у вигляді дрібних кулястих лейкопластов, що групуються навколо клітинного ядра.
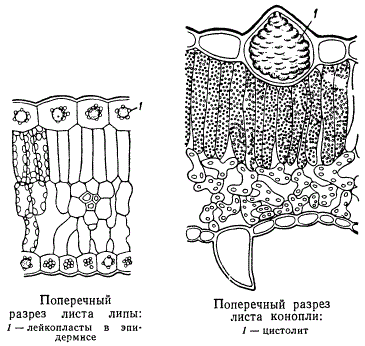
У епідермальних клітинах утворюються різні продукти життєдіяльності протопласта. Особливо своєрідні цістоліти. , Вони представляють собою досить об'ємні відкладення вуглекислого вапна, що виникають в гіпертрофічна розрослися клітинах епідермісу (ідіобласти, в даному випадку - літоцістах). Якщо подіяти на ці цістоліти кислотою, наприклад, соляної, то основна маса цістоліта розчиниться і залишиться лише целюлозна основа, прикріплена до оболонки клітини, на якій відкладається вуглекислий кальцій. Форма і розміри цістолітов у різних рослин різні. Вони властиві певним видам рослин. У нашому прикладі наводиться цістоліт в листі конопель. Класичним прикладом служать цістоліти в листі фікуса.
Оболонка клітин епідермісу, особливо зовнішня стінка їх, часто просочується або солями кальцію, або сполуками кремнію (злаки, хвощі). На поверхні епідермальних клітин дуже часто виділяються кутикула і кутикулярного шари. Кутикула затягує всю поверхню сформованого епідермісу, перериваючись тільки устьічнимі щілинами. Крім кутикули, зовні її у більшості рослин виділяється віск у вигляді тонкого дрібнозернистого нальоту. У деяких тропічних рослин восковий покрив на листі досягає значної товщини (0,5 см). І кутикула і восковий наліт не змочуються водою і оберігають тканини, розташовані під ними, від втрати води.
Умови зростання визначають деякі особливості структури епідермальних тканин рослини. Епідерміс листя і стебел водних рослин майже не має кутикули і тим більше воскового нальоту, а у рослин посушливих місцезростань кутикула, як правило, утворюється. Оболонки епідермальних клітин надземних органів деяких рослин дерев'яніють, стають вельми товстими, з сильно скоротилися розмірами клітинних порожнин. При цьому часто дерев'яніють не тільки клітини самого епідермісу, а й підстилають його клітини. Поверхня епідермісу надземних органів дуже рідко залишається гладкою і однорідної протягом всього часу існування цієї тканини. Навіть на кутикуле майже завжди виникають горбки, складки, штрихи, які надають поверхні різний, іноді досить примхливий малюнок. Крім того, клітини епідермісу часто утворюють найрізноманітніші вирости - волоски. Волоски бувають одноклітинними і багатоклітинними, гіллястими і неветвістимі. Форма волосків також варіює: існують волоски зірчасті, головчатиє, у вигляді лусочок, пріцепок і ін. Форма, розміри і розподіл волосків на поверхні органів характерні для певних груп рослин, і ці ознаки часто служать систематичними.
Зазвичай волоски відокремлюються перегородкою від тієї епідермальній клітини, на якій вони утворилися. Але є волоски, що представляють собою навіть у цілком розвиненому стані не більше як вирости клітин епідермісу, що не відокремлюються від них спеціальної перегородкою. Якщо такі вирости епідермальних клітин дуже короткі, то їх називають сосочками. Сосочки особливо поширені на пелюстках квіток, надаючи їм бархатистість.
Багатоклітинні волоски нерідко надзвичайно сильно розростаються, складові ж їх клітини дерев'яніють і волоски стають твердими. Такі волоски називають шипами(Малина, старі стебла гарбуза, стебла деяких сортів троянди, барбарису і ін.).
Все різноманіття волосків, керуючись фізіологічним принципом, можна звести до двох основних типів: криютьі залізисті. Дуже часто волоски обох типів існують на одному і тому ж органі. Наприклад, на малюнку зображено прості криють і залізисті волоски приквіткове лусочки рудбекии.
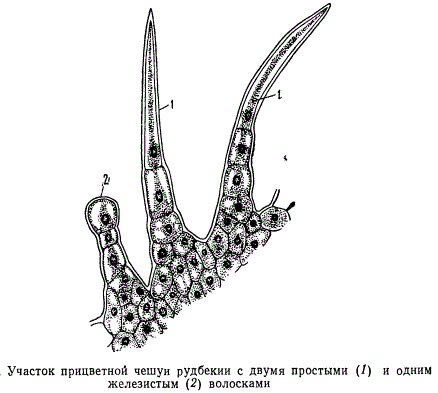
Кроющие волоски залишаються живими порівняно недовго. Коли протопласт волоска відмирає, остов його, що складається з оболонки, зберігається, наповнюючись повітрям. Зазвичай ці наповнені повітрям блискучі білі кістяки волосків і утворюють такий характерний волосяний покрив тіла рослин.
Морфологічна різноманітність криють волосків представлено на малюнку, на якому зображені волоски частин квітки рослин з сімейства норичникових. Всі волоски за винятком двох одноклітинні.
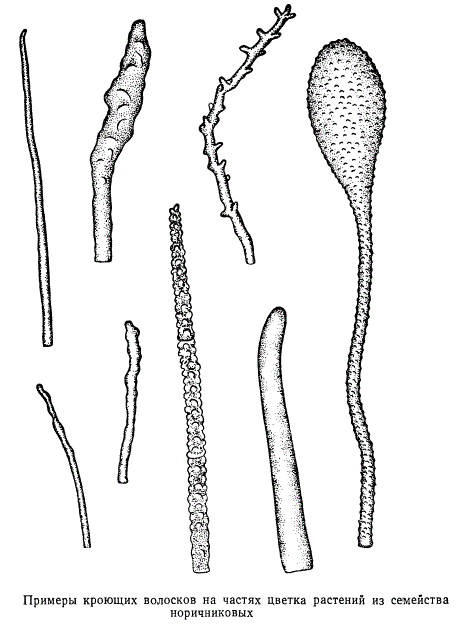
Залізисті волоски зберігають живе вміст своїх клітин трохи довше криють. Протопласт клітин залізистих волосків, особливо верхівкового, що утворюють головку волоска клітин, нерідко виділяється значною вакуолізацією і майже завжди містить велике клітинне ядро. Якщо протопласт клітин залозистого волоска руйнується, то ніжні оболонки їх спадаються і волосок, засохнув, відпадає, не залишаючи після себе майже ніякого сліду. Морфологія залізистих волосків так само, як і криють, безмежно різноманітна. Зазвичай виділення залізистих волосків складаються або з смолистих речовин, або ж з ефірних масел, цукристих речовин, слизу, а іноді просто з води і розчинених в ній речовин різної природи.
Залізисті волоски, що покривають, наприклад, ниркові лусочки і криють листи нирок, виділяють смолиста речовина, як би закупорють нирку і охороняє ніжні листочки від висихання. Можливо, що виділення залізистих волосків є кінцевими продуктами складних біохімічних процесів, що протікають в рослинних організмах.
Залізисті волоски, що виділяють воду і мають спеціально пристосованої для цього структуру, називають гідатоди.
Серед залізистих волосків особливо своєрідні за своєю організацією так звані пекучі волоски, загальновідомим прикладом яких є пекучі волоски кропиви. Стінки волоска кропиви просякнуті сполуками кремнію, внаслідок чого стають дуже ламкими, особливо у верхівки. Такий волосок обламується від найлегшого дотику, причому кінчик волосини перетворюється в гостру голку, легко встромлюють в шкіру тварини або людини. Такі ж волоски з ще більш пекучим секретом утворюються на різних частинах Alonsoa. У більшості інших випадків оболонки залізистих волосків ніжні, навіть якщо вони і виділяють дратівливий шкіру секрет, як це має місце, наприклад, у деяких видів борщівника.
Значення волосків різноманітно і в багатьох випадках ще не з'ясовано. Прийнято вважати, що найбільш поширеною функцією волосків типу криють є запобігання поверхні тіла рослини від зайвої втрати води. Блискучі білі волоски, з одного боку, відображають прямі сонячні промені, з іншого - густим сплетінням створюють над епідермісом середовище, яке сприятиме зменшенню транспірації. Однак спеціальними дослідами принаймні щодо частин квітки показано, що покривають їх життєдіяльні волоски збільшують інтенсивність транспірації.
Зазвичай ознайомлення з різними волосками засноване на вивченні волосків, що покривають вже виросли органи. За винятком залізистих волосків, описуються волоски мертві, позбавлені живого вмісту. У живому стані такі волоски, безсумнівно, більш пов'язані з процесами життєдіяльності рослинного організму і грають іншу фізіологічну роль, ніж відмерлі сухі волоски. Підтвердженням цього положення є розвиток волосків на колосках злаків. На молодих квіткових і колоскових чешуях колоса, коли він ще перебуває в стадії так званої трубки і остаточно ще не оформився, по краях лусок, уздовж середньої жилки і на молодий ості, якщо колос буде остистим, розташовані волоски з добре вираженим великим клітинним ядром і густий цитоплазмою. Ці волоски перебувають в найбільш активному життєвому стані. З усіх частин колоска колоскова і квіткова луски розвиваються в першу чергу, а волоски на них формуються раніше всіх інших клітин, складових тканини цих лусок. Волоски на чешуях колоска залишаються в діяльному стані в усі час формування зав'язі і зародкового мішка в ній. Як тільки зародковий мішок готовий до запліднення або, в інших випадках, коли запліднення відбудеться, вміст волосків починає швидко відмирати. Волоски, розташовані уздовж середньої жилки і ості, встигли значно розростися, перетворюються в шипики і горбки, зазнають одревеснение і надають колосу добре відому на дотик шорсткість.
Таким чином, волоски на починаючому розвиватися колоску злаку в свою діяльну стані пов'язані з найважливішими процесами в житті рослини - формуванням і дозріванням статевого апарату, під час яких йде найактивніший обмін речовин.
Форма волосків на молодих колоскових чешуях різноманітна.
На молодих зав'язях різних рослин часто утворюється багато волосків, у вмісті яких є алкалоїд або будь-яке інше фізіологічно активна речовина. Всі ці волоски в період формування зав'язі знаходяться в життєдіяльному стані, а після запліднення починають відмирати і відпадають. На малюнку зображено залізисті волоски зав'язі оману; пізніше, після запліднення, вони швидко зникають. Ясно, що волоски, настільки добре і в достатку, що розвиваються на поверхні зав'язі і відпадають після запліднення, в своєму розвитку пов'язані з інтенсивно йдуть в зав'язі процесами формування жіночих статевих елементів.
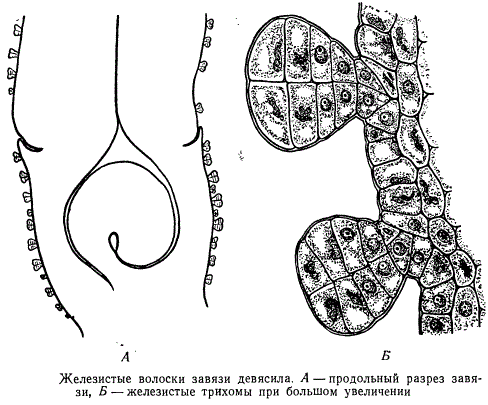
На сім'янку однією з ромашок, які ростуть в пустелі, можна знайти своєрідні волоски, зображені на малюнку. Кожен волосок складається з двох морфологічно добре відрізняються частин: підстави, що складається з декількох члеників - окремих клітин, і верхівки, зайнятої однією ослизнюються кліткою. Кожен членик підстави забезпечений целюлозними тяжами потовщень оболонки, що утворюють витончений палітурка - подобу каркаса. На малюнку всі три волосини представляють собою різні стадії ослизнення кінцевої клітини. Ослизнюються клітини і волоски на поверхні плодів деяких рослин, які ростуть в пустельних місцевостях, відокремлюються відносно дуже рано, вони добре помітні на далеко ще не зрілих плодах. Часто такі клітини у вигляді коротких товстих волоскоподобних утворень розташовуються на верхівках ребер семянок, причому при посередництві особливих клітин, які називаються гідроцітамі, Оболонки яких забезпечені різноманітними стовщеннями, зовнішні волоскоподобние клітини повідомляються з судинними пучками.
Таким чином, створюється своєрідна гідроцітная система, яка відіграє велику роль у водному режимі дозріває плода.
За дозріванні семянки волоскоподобние гігроскопічні освіти засихають і залишаються на поверхні у вигляді невеликих горбків і бородавочек або опадають, або ж перетворюються в ослизнюються клітини.

Епідерміс утворюється не тільки зовні органу, а й усередині, якщо в органі формується спеціальна порожнину. Наприклад, тканина, що вистилає порожнину плодів, за своїм походженням ідентична епідермісу, що покриває плоди зовні. У зв'язку з перебуванням покривної тканини всередині порожнини, де вологість повітря та інші умови відрізняються від умов зовнішнього середовища, особливості структури внутрішнього епідермісу плода дещо інші, ніж зовнішнього епідермісу.
Клітини внутрішнього епідермісу плода гороху з'єднані між собою пухко, кутикула або дуже тонка або відсутній. Значна частина клітин витягнута в волоски різної довжини з тонкими оболонками. Місцями і особливо близько насіння утворюється густий пушок з сосочкоподобние волосків, огортав насіння з усіх боків. Те ж спостерігається в плодах кінських бобів, тільки покрив, забезпечений волосками, відрізняється більш густим і рівномірним скупченням останніх.
Безсумнівно, освіта таких сосочкоподобние волосків, що стикаються з країнами, що розвиваються насінням, має якесь фізіологічне значення в процесі розвитку насіння.
У деяких рослин оболонки внутрішнього епідермісу плода досить значно товщають і іноді дерев'яніють, особливо на часі остаточного дозрівання плоду. Хорошим прикладом такої будови внутрішнього епідермісу плода є епідерміс, що покриває зсередини стінки коробочок маку. Волоски в таких випадках розвиваються не завжди і ніколи не досягають значної довжини; кутикула виражена більш помітно, поверхня епідермісу часто буває гладкою, клітини мають потовщені оболонки і щільно зімкнуті між собою, іноді серед них зустрічаються продихи.
Таким чином, роль волосків і подібних до них утворень у житті рослинних організміввельми різноманітна і змінюється в залежності від умов зростання і віку рослин.
Тривалість існування епідермісу на надземних пагонах невелика і зазвичай обмежується одним вегетаційним періодом. Але є винятки і не настільки рідкісні, особливо у рослин більш південних районів. Наприклад, листя лавровишні і лавра існують більше одного року, отже, і епідерміс їх зберігається більше одного року. Є дерева, у яких епідерміс на пагонах зберігається кілька років, навіть на товстих стовбурах багаторічних дерев. Прикладами служать евкаліпт, платан, стеркулія, а на півночі - сіра вільха, черемха і ін. Многоліття епідерміс також на хвої сосни, ялини, туї та ін.
На поверхні багаторічних епідермісу розвивається щодо товста кутикула. У хвойних оболонки епідермальних клітин дерев'яніють.
На відміну від надземних частин рослин тканина, що покриває молоді частини коренів, позбавлена кутикули внаслідок чого клітини цієї тканини мають оболонки, легко проникні для води. Крім того, у більшості рослин при відповідних умовах клітини, що покривають кінцеві ділянки коренів, утворюють тонкі вирости, що не відокремлюються від утворили їх клітин перегородками. Ці клітини називаються кореневими волосками. кореневі волоскиу різних рослин бувають різної довжини, але в загальному довжина їх коливається від 0,15 до 8,0 мм. На коренях проростків гороху, вирощених у вологому атмосфері, довжина кореневих волосків дорівнює 2,5 мм, у кінських бобів - 0,8 мм. У одного і того ж рослини в залежності від умов середовища, в якій розвиваються коріння, довжина кореневих волосків може значно коливатися. У більш сухих умовах кореневих волосків утворюється більше, ніж у вологих; у рослин, які ростуть в грунті, покритої водою, кореневі волоски часто абсолютно не утворюються.
Оболонка кореневих волосків целюлозна, дуже ніжна, кутикула на життєздатних волосках не утворюється. Найбільш звичайна циліндрична форма кореневих волосків. Гіллясті кореневі волоски формуються лише у рідкісних рослин, але при склеюванні з частинками грунту строгість циліндричної форми їх порушується. По суті, кореневі волоски представляють собою вирости клітин епідермісу, нерідко сильно подовжені. Однак у деяких рослин кореневі волоски все ж відокремлюються перегородкою від породила їх епідермальній клітини.
Починають розвиватися кореневі волоски не на самому кінчику кореня, а на деякій відстані від нього, приблизно на 0,5-3,0 мм від верхівки. Зона, яку займає кореневими волосками, по довжині кореня не дуже велика, причому довжина зони залежить від багатьох умов, але не перевищує декількох міліметрів. Кореневі волоски недовговічні: досягнувши певної межі існування, вони відмирають, зморщуються і відпадають. Відмирання кореневих волосків відбувається в тому кінці займаної ними зони, де розташовані найбільш старі волоски, а на протилежному полюсі зони відбувається новоутворення волосків. Кореневі волоски постійно виникають знову в більш молодих ділянках кінчика кореня і відмирають в старіших, структура яких стає більш постійною. Таким чином, розгалуженнями кореня охоплюються всі нові і нові частинки грунту, з якими злипаються кореневі волоски. Кореневі волоски розташовані на кінчиках коренів в займаній ними зоні дуже густо. Наприклад, у кукурудзи на 1 мм 2 поверхні кінчика корінця припадає близько 425 кореневих волосків, а у гороху при тих же самих умовах - 232. За рахунок утворення кореневих волосків поверхню кореня сильно збільшується. Так, поверхня покрита кореневими волосками частини кінчика кореня кукурудзи в 5,5 раз більше поверхні такої ж частини кореня без волосків, а у гороху - навіть в 12,2 рази. Збільшення поверхні кореня сприяє виконанню однієї з основних його функцій - поглинання води і розчинених в ній речовин.
Якщо ви знайшли помилку, будь ласка, виділіть фрагмент тексту і натисніть Ctrl + Enter.