Който има клитинна стинка. Будова и животът на актиномите
прокариотна клитина
прокариоти- организми, но не вода, на базата на еукари образуваме клетъчно ядро и органоиди на вътрешната мембрана (зад винетка от плоски цистерни във фотосинтетични видове, например в цианобактерии). Сам по себе си е велика килцева (при някои видове-линея) двулантюжкова ДНК молекула, в която основната част от генетичния материал на клетката (т.нар. Нуклеоиди) не подкрепя комплекса с хистонови протеини (т.нар. Хроматин) . Бактерии, включително цианобактерии (синьо-зелени водорасли) и археи, се въвеждат преди прокариотите. Местата на прокариотни клетки са органелите на еукариотни клетки - митохондрии и пластиди. По принцип, вместо клетките, които ще запомнят цялата й obsyag, има свързване на гранулирана цитоплазма.
еукариотна клитина
еукариоти- организми, които са Володими, на базата на прокариоти, ние ще образуваме клетъчно ядро, отделено от цитоплазмата от ядрената обвивка. Генетичният материал на гънките в децилинейните двуверижни молекули на ДНК (под формата на комплекс от организми, броят на ядрото може да варира от две до няколкостотин), прикрепен от средата към мембраната на клетъчното ядро и възстановен , редици хроматин. В клетките на еукарот е система от вътрешни мембрани, които фиксират ядрото на ядрото, редица други органоиди (ендоплазмено сито, апарат на Голджи и др.). Освен това в най -важния и постоянен вътрешен симбионт -прокариоцит - митохондриите, а във водораслите и рослина - също пластиди.
2) цитоплазма
цитоплазма- вътрешната среда на жива или мъртва клетка, граничеща с ядрото и вакуолата, е заобиколена от мембрана. Тя включва хиалоплазмата, която е основното проникване на цитоплазмата, която се намира в нео -лигаментните клетъчни компоненти - органели, както и развитието на непотентни структури - включването.
Всички видове органични и неорганични речи... В тяхно присъствие също липсват комуникационни процеси и резервна реч. Основната река на цитоплазмата е водата.
Цитоплазмата постепенно се срутва, прелива в средата на жива клетка, променяйки едновременно със себе си развитието на речта, включването и органелите. Tsey rukh се нарича циклози. Те имат всички процеси на обмен на думи.
Цитоплазмата е здрава, докато расте и расте, а когато е частично видима, може да бъде подновена. Нормалното функциониране на цитоплазмата обаче е само в присъствието на ядрото. Цитоплазмата не може да бъде без ново развитие, така че самото ядро е без цитоплазма.
Ролята на цитоплазмата на полето във формирането на всички клетъчни структури (компоненти) и в поддържането на химични взаимодействия. Така че самата цитоплазма подстригува тургор (обсяг) клитини, температура на отклонение.
3) Клитинна стинка
клитинна стинка- твърда обвивка от клитини, изпечена от цитоплазмената мембрана и структурата, транспортните и транспортните функции. Vyavlyayut на големи бактерии, археи, гъби и roslin. Същества и най -прости неща духовна история.
Слитни стинки прокариот
Класът бактерии се съхранява в пептидогликан (муреин) и има два вида: грам-положителни и грам-отрицателни. Клетъчният тип на грам-положителен тип се съхранява в една топка пептидогликан, която се прилепва към клетъчната мембрана и прониква с тейхоевия и липотейхоеви киселини. При грам-отрицателни видове топката пептидогликан е по-тънка, между нея и плазмената мембрана има периплазма, а задната част на клитина се излъчва от друга единична мембрана, представена от Т.
Слитни стинки мушив
Класическите гъби се съхраняват в хитин и глюкани.
Клинична стинка.Това е багатошаров корпус с 9 ... 10 топки от електронната индустрия. Система от микрофибрили, вградени в аморфна матрица, оформена като скелет на челини. Влакната в утайката на вида могат да бъдат сгънати от целулоза, глюкон и хитин. Инши полизахариди, билки, пигменти, липиди служат като цимент с думи, весели връзкиот микрофибриларната част на класа. Появата на такива комплекси ще предотврати вибрациите на проникване за някои речи и блокадата на други.
Подкрепящите микроби и клиничните принципи и матрица се основават на механизма на образование и биосинтез. Фибрилите и матрицата се възстановяват асинхронно, а фибриларният скелет на стъблото се регенерира на първо място. Биосинтезата на две части от културата на света е включена в участието на ензими.
Процесът на установяване на политика се показва по два начина: нов материалможете или да се vprovadzhuvatsya в полярността на стената, или еднакво да се припокриват по цялата повърхност. Първият вижда одобрението на цилиндрични клетки, другият - сферични.
Клинична стинка да служи като постоянна привързаност и да обединява гъбен съединител от притока на млади служители в средата на колонията, например осмотична лента, която ще обобщи вибрационното проникване за млади речи. Вона даде форма на вегетативни клетки от хифи и репродуктивни органи. На повърхността на клетъчната линия и цитоплазмената мембрана на локализирани ензими, както и трансформация на полимерни материали, които не се използват от клетките (не се намират във вода).
В резултат на лизиса на клетките гъбите могат да се поръсят с инфузия на ензими, които могат да се разглеждат като малки клетки и да се установят в клетките на самата гъба.
Основните компоненти на клетъчната линия на гъбичките са хитин, глюкан, протеини и мазнини. Азот и безазотни полизахариди с мастни гъби могат да се използват за създаване на различни и несъществени комплекси. 4 ... 6 монозахариди формират основата на духовния стил; Глюкозамин, маноза, глюкоза, ксилоза и др. Са включени в склада на полизахаридни фракции. Плъзгащият се пикрес, който е складът на канелената обвивка на младите клитини от същата гъба, не е същият.
протопласт- вместо клетки, поставени в клетъчната стена: МА цитоплазмена мембрана, ендоплазмен ретикулум, едно или повече ядра от ядрото, както и митохондрии, рибозоми от РНК, лизозомия, апарати на Goldzhin, вакуоцити, вакуоцити и включени.
Цитоплазмена мембрана.Тънка тришарова черупка, се отделя без средата от клетъчната стена и от цитоплазмата. Цитоплазмената мембрана прониква вибрационно за говор, така че да може да влезе в клетката и да излезе от нея. Цитоплазмената мембрана отмъщава до 40% липиди и до 38% липиди. Ризната форма на инвагинация и обменът на цитоплазмената мембрана се наричат мезосомия.
Основната функционална характеристика на цитоплазмената мембрана на полето при настъплението е: добро здраве в клетката на младата реч, ензимна обработка и появата на продукти в метаболизма. Реорганизира се в цитоплазмената мембрана на речевата линия, за да влезе в протопласта на клетъчната линия и да участва в обмена на речеви линии.
Ендоплазмения ретикулум.Съхранява се от луковици, тубули и вакуоли, услуги до безплатни депа на оживени реки.
Митохондрии.Числено разпадащи се затворени капаци с елипсовидна форма, с прегради, покритие с една или две топки. Признавайки, scho митохондрии, установяването на собствената ДНК на пръстеновата структура, изградена преди възпроизвеждането. Митохондриите са отохенизирани с мембрана, върху която се наблюдава локализацията на ензими: пируват оксидаза, фосфатази с янтарна киселина, пероксидаза и други ензими. Митохондриите служат като генератори на енергия в клитина. Формата на митохондриите и броят на сортовете в класа винаги е въпрос на умове за култивиране и физиологичен статус на клиентелата.
Рибосоми.Заоблени зърна с рибонуклеотидна природа с размер до 200 \ участват в синтеза на клетки. Броят на рибозомите е значителен за растеж при по -старите видове гъби и застой от различни фактори, в културата и вътре.
Апарат Голджи.Представяне на група крушки с дъгообразен диаметър (0,000 002 ... O, TOV 01 микрона) или успоредни на лежащи дисковидни плочи. Целият органоид расте в клетки на дилент, под формата на рибозоми.
Лизосоми.Те отиват в апарата на Голджи, разбъркват се между клетъчната мембрана и цитоплазмената мембрана. Това е гранулирано осветление, оточени едносферична липопротеинова мембрана. Отмъстете за ензима, хидролизирайте блоковете и действайте като рецептор за клитина като неприятен приток на токсични думи, екзо- и ендогенен поход.
Липосоми.Капки мастни потоци, оточени мембрани с една топка.
Ядро.Намира се в центъра или на полюсите на клитини. Гъбичните клетки могат да имат единични и множество ядра. Вонята сякаш стои зад спада на функциите. Формата на ядрата е закръглена и подовжена. Дермалното ядро е облицовано с двусферична пореста нуклеомембрана с ядро от твърди зърна и тънки фибрили. Ядрото трябва да се намери в склада на ДНК хромозомите. Чрез анастомозите на ядрото можете да преминете от една клетка към инша.
Включване.В гъбните клетки са включени следното: волутин, гликоген, липиди, пигменти, милоидно здраве, соли на органични киселини, аминокиселини и други. Вважае се, че гликогенът води до ендогенно поведение, а волутинът служи като резервна животворяща реч, тъй като участва в енергийните процеси.
Това означава, че в процеса на живот в клетките на гъбичките се натрупват различни метаболитни продукти - антибиотици, ензими, токсини, витамини и други.
Всички числени морфологични елементи на микроскопичните гъби са разделени на две групи: мицели и спорове. Вонята бръмчи с розови форми и форми. Морфологичната функция на супержилата и мицелата е да служи като важен диференциален символ за вида гъбички.
Мицелий.Това е кръгла тръба vuzka, чийто диаметър варира от един микрон до децилен микрон.
С ryazhenní gífі mіceliyu, стикайте един по един, можете да зададете клиентите на zlittya mízh, - анастомози. С проявата на голям брой мицели присъства типичен виглиад. Развитието на анастомози се насърчава при млади гъбички от големи клетки. Животновъдите могат да прехвърлят клетъчното ядро от една клетка в една клетка и да преминат от хаплоидна към диплоидна мицела. Въпреки това, при голямо разнообразие от воня, вегетативните функции се развиват и развиват във форми на багатеч с недостиг на храна. Довжина клитин mycelіyu може да се използва от decílkoh микрони до десетки и повече от стотици микрони.
Разряден мицел с двуконтурна обвивка, като в млади култури на нижниша. При преградите, където към краищата на клетките се добавя мицелия, цитоплазмата се излива през тях в процеса на растеж и от нея се получава оживен говор. В клетките на голям брой млади, цитоплазмата става гранулирана чрез вакуолите при възрастните хора. Младият мецелий се съхранява от подвженни правоъгълни клетки, старите - от къси заоблени или с големи фасети. Мицелът, който има преграда, се нарича пресечен. В случая с долните гъби, мицелите се съхраняват от хифите, които са отстранени от напречните прегради, и е като би една, силно заоблена гигантска клетка с числени ядра и се нарича несептирани мицели.
Как виждате развитието на мицелия? Три свръхпотока випинат Паросткова тръба, като подовжуват се и след това се отделят с преграда от средната част, която включва спор. Растежните тубули ще растат все по -леко и ще получат нова преграда, разпределена към дисталната, горната, клетъчната и проксималната, или вътрешната. Nadal verhhivkova клетка podovzhutsya и знаят дълго, вижте приятел, млад мъж на първо място, вътрешна клетка. Така че се повтаря и дава. Целият процес на вътрешните клетки само расте, напречно нараства леко, но след това те развиват малки глави. В дисталния край на вътрешната клетка има малко випиня, която приема цилиндрична форма и се появява зад преградата като виробля на клетката. Нова клитина израства в малка бутилка, разпростира се и се разклонява в същия ранг, като глава. Ръководителите на развитието на гилок с глава, простираща се от главата смърди, са по -стари и по -силни от тези, които са по -близо до основата, че външният им вид е акропетално.
Развитието на несептифицирания мицел се показва в загалния в същия ранг, макар и без одобрението на напречните прегради. Растежът нараства върху нишките на хифата, деакумулираща протоплазма, която ще запомни цялото образование, а в по-големите задни части има значително развитие на централните вакуоли. В същия среден клас, например, на повърхността на животворния желатин, мицелът на хифи (както неклетъчен, така и неклетъчен) расте еднакво и радиално, така че мицелът расте под формата на кола, тъй като расте от ръбовете. Централната част в най -новата е най -новата, а периферната част е най -новата.
В случай на загално монотонно развитие на мицела, което може да се нарече типично, в близост до есента се насърчават редица специфични оризове, като макроскопичния вид и захалския характер на растежа, както и микроскопичните будови. Макроскопски изглед на мицелия е показан от дясната страна на екрана. При някои капчици вонята се образува върху самата повърхност на субстрата и частично в средата му и само в мицела на плоска виглиада, притисната до субстрата на ръба; при някои видове, освен това, все повече хора растат, така че те да се появяват всеки ден и да приличат на мицелия, например, с парче вата, така че да се издига над субстрата. Характерът на растежа може да бъде различен при една и съща гъба в състояние на упадък, прибиране на реколтата и в. Въпреки това, редица форми на гъби имат специфични характеристики, например установяването на писмена хранителна гъба - руина на дърво.
Броят на мицелите е най -често срещан в средата на деня, ейл от върха на кафявия връх на зимния сезон. Цената е обвързана с прикачването на програмата в клиентелата и по -рано в средата на самия клитин.
Развитие на истински и псевдоклетъчни клетки. Остава да се характеризира с факта, че около клетките те не са свързани една към една и не губят външната обвивка. Заместването на справедливото разпределение тук е да се насърчи развитието на дървовидния растеж на клитина.
За прикрепване към субстрата и използване на оживени речи в хода на еволюцията на гъбите бяха формулирани специални индикации за цялата организация: Ризоидите са с къса коренообразна форма, а апресорите са къси продължения, а някои са с форма на лопатки и вирусна мицелия.
склероция, Тежки, ризоморфни и хламидоспори също са видове микроклетъчен растеж.
Склероци е отделяне на гъбични хифи, които го правят специален. Когато се образува склероция, мембраните на хифите трябва да се запълнят по -тъмно. Склеротиите на хифите се насърчават силно, в средата на хифите има повече тънкости и не са фантазии. Склероцити - цената на предварително напрегната храна, която позволява на гъбата да прекара тривиален час в разгара на лятото и да попречи на нейното израстване до притока на по -нови фактори: температура, сънливост и индустриални условия. Развитите склероции трябва да бъдат заменени с по -малко vologues по отношение на микроклетки и много резервна реч - липиди, гликоген.
Размерът на склероциите варира от децилни милиметри до децили десетки сантиметри, а формата на склероцията е сферична, неправилна;
Структурата на клетъчните склероции и механизмът за определяне на растежа, образуването на протеини в процеса на подобряване на разпределението на мицела и отделяне на хифи. Има два начина за възпитание на склероции: термичен - в края на хифата; интеркален - във външните фрагменти на цефалните хифи.
В гъбите багаток, с развитието на овощни дървета и определени вегетативни структури, се установява помилкова тъкан - псевдопаренхим (псевдопаренхим). Въз основа на паренхим от дясната тъкан, в резултат на винарната, има три прави линии, плектенхим се преструват на клюки и растеж. Ако трябва да го съхранявате от клитиновите големи-изометрични изометрични, тогава тя се нарича параплектомия; Ако в най-добрия момент е ясно, че глифоподобната Будова (клитини с форма на подовжени), тогава тя се нарича прозоплектенчимой.
тежки микроклетъчни- вегетативна структура на линейно агрегирана хифа. Диаметърът на микроклетъчните нишки се отлага като редица геофони, които са концентрирани близо до централната основа.
За най -простия тип няма голям брой паралелни водачи, които да бъдат залепени една към една бавно наречени черупки или да се влезе в по -голяма машина с път на коефициенти. муване на числени кратки анастомози. На първо място. В чайовете, ако са тежки, те ще получат особена разлика. Имената на елементите са по-тънки, което го прави като би-кора близо до централния товстов стовбур.
Ризоморфи- по -сгъваема в агрегацията на hyfi, която расте при интензивния растеж на централните хифи в растящите гъби, дължината на хифите, а също и от етапа на диференциация на клиентите на хифата.
Името на частта от ризоморфа има тъмен цвят и подобно сходство с корените виших рослин... Вонята е широко разпространена в гъби с големи плодови тонове: в базидиална, сумирана и в.
По принцип характеристиката на микроклетъчните нишки и ризоморфния полиагус е в запазването на експандираните гъбички в субстрата и свръхпредлагането според хифите на старата реч.
боклук- цената на промяната на мицела при възрастни и стари култури при деца, или по пътя. Основната функция на хламидоспорите не е разпространението, а запазването на ума. Формата на техните охлюви е кръгла, овална или леко повдигната, диаметърът се променя над диаметъра на мицела. При някои гъби стената е двойно контурна, повърхността е гладка или къса. Chlamydospore може да се намери по краищата на мицела, така че вонята се нарича термична, по пътя - интерполярна (индустриална).
По -старите култури често поддържат голяма колекция от хламидоспори от химерната форма, което е малко анормално. Млади и зрели отпадъчни спори за покълване. Старите клетки са дегенерирани.
спорят... За допълнителна помощ спорите на гъбичките не се размножават, но също така се разширяват в средата на колонията. Тесен поток от временността на обвивките на свръхпотока към притока на агресивни бюрократи. Спорите за ендоспори, където са установени всички специални обекти - споранги (торби), екзоспори, розташовани върху микроскопа.
При младите гъбички те спорят срещу ооспори, зигоспори, аскоспори, базидоспори, ендоспори, фиалоспори, хламидоспори. Оспорват непълните гъби в зависимост от размера на групата. Преди Ендоспора е необходимо да се установи в средата на пътя на сегментирането на останалите, да носи талоспори, които включват артроспори, хламидоспори и бластоспори. Нещо повече, за маломерните гъби е характерно да установяват конидии, макроконидии, алеирии (микроконидии) и полукълба, в които доминират маломерните конидии.
Хемиспорията е по -леко свързана с мицела и представлява един или два сегмента, за да бъде развързана за напречната дължина на мицелната нишка. Формата им е цилиндрична, техните са закръглени или с големи фасети, черупката е двойно контурна.
За доставка на голяма част от масата сухи гъби в склада, и от 5 до 15%. Складът е много променлив и често е много специфичен за пеене на таксономични групи гъби. Цената може да бъде закупена от склада в склада от представителите на старите, хитридни и мукозни гъби - зигомицети, посочени в таблицата. 1.3 (Аронсън, 1965).
* (Количество пепел без фосфати.)
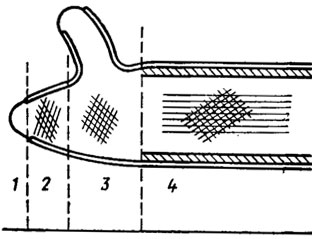
Структурно черупките на гъбичките се стимулират на базата на двуфазна система, в която микрофибрилите са включени в аморфната матрица. За данните от електронната микроскопия има не по -малко от две топки в просто, ясно оформление на фибрили. Вътрешната топка започва да се движи по оста на главата на клетката и призивът е да слезете до нея (фиг. 1.1). В други черупката е малко багатошаров, освен това мананът е локализиран в най -често срещаната сфера, а глюканът е локализиран във вътрешната тънка. Във водни гъби, например, в Allomyces, черупката е направена от псевдосепти - псевдосептици, които могат да направят спиците на колелото. При обикновените и базидиалните гъбички спонтанните прегради - септи - са по -чести. В случай на призоваване на септи миж клитини, може да искате едно просто време, при нарушение на страните на което в процеса на формиране е възможно да се бачи върху двойката осмофилни деца на Воронин. При базидиалните гъби на пори те често са още по -сгъваеми, несигурни ковпачи - родители (фиг. 1.2). Такива пори були са известни в овощни дървета и ризоморфи в редица други базидиомицети (Moore, 1965; Burnett, 1968). Въпреки това, до настоящия час не е необходимо да има ум, който да не е податлив на естествени промени в структурата на септалните върхове на аско- и базидиомицетите към тяхната таксономична привързаност, дори към хетлоидната сума на преобладаващите хомокаротични структури. Такъв doslіdzhen е умиращ, ale ihkh значение за еволюционните и таксономични импулси в царството на гъбите по същия принцип.
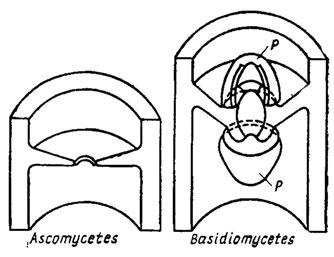
Bagato new беше добавен към оживяването на септалните гъби до края на деня на електронния микроскопичен етап (Flegler et al., 1976; Kamaletdinova, Vasil'ev, 1982). На първо място, той се превърна в своеобразен делфин от базидиални гъбични структури, мабут, херметично изолиращи клетки, мицел от една до чак до кочана на овощни дървета (Flegler et al., 1976). Процесът на изолиране е успешен, за да бъде разпознат от протеолитични ензими и незабавно осмофилен (да замени бутилките и липидите) с двустранни тапи, които са известни до момента на образуване на плода. Їх се актуализират знанията за наблюдение чрез перфорации с родители и получаване между клетки на глифове.
Оставайки внимателни за преградите в плодните тела на дискомицети (например Peziza badia - Kamaletdinova, Vasil'ev, 1982) показа, че е по -често клетките да растат структурирани в класа на обикновените гъби. В онези, които се формират, те поемат съдбата на таланта на Воронин, за да се утвърдят в иновационната форма на колбата на клетъчната мембрана, осмофилна на мястото на тези (талантът на Воронин), за да могат да преминат през преградата и да се обърнат далеч За по -нататъшно проникване на Tyltsya Voronin в аномалиите на хименната топка, мабут, превишаване, специална структура е перфорирана, така че човек може да лежи повече от час в клетката на майката на майбутната торба, тъй като изглежда, че прониква във вътрешните органи на органа. Някои от неразтворимите структури се насърчават и оформят в плодните тела на парафините.
Допълнителни структури в спороносните мицели на непълни гъби, за да отмъстят за наклоните на Воронин, например, от Arthrobotrys conoides. При производителя на пеницилин дейтеромицети Penicillium chrysogenum структурата се разкрива в септиците, точно както е известно от Камалетдинова и Василиев в дискомицетите Peziza badia (Kurilowich et al., 1980).
Скелетните основи на черупките на гъбите се отлагат от кристални органични полизахариди: целулоза, хитин, хитозан, манани, глюкани и иншим. Цялата воня може да има линейна структура с β-1,4-връзки на външните компоненти-мономери на хексози, амино и ацетаминохексози. Според резултатите от микрохимичното тесто (предварително прибран Rutenium chervonim), пектинът е бил предварително въведен в складовете на черупките на гъбите. Резултатите от химическия анализ обаче не потвърждават наличието на мономерен пектин галактуронова киселина преди структурата на пектина в склада (Aronson, 1965).
Хитинът и хитозанът за голяма част от гъбите са още по-характерни в складовете на черупки, като азотсъдържащи полимери. С голям хитин от гъби, той е още по-подобен на кома и ракоподобен хитин, което се потвърждава от картина на тяхната дифракция в рентгеновите обмени. Азотът в гъбите обаче е по -малък от азота, по -малко в близнаците и в средата складови части Bula viyavlena метилпентоза, наречена мицетоза. Хитинът в гъбите може да се намери в Microchimichny по метода на Van Wisseling, зад добавянето на малка локва деацетилиране и обидна реакция към хитозан и чрез дифракция на X-промени. Спечелете, като не знаете само в оомицети, като гъби Saprolegnivi и Peronosporovin. По -рано беше важно нормално другите хора да останат в клетъчните стени - захарните прегради (Кулаев, 1975).
В датския час се установява, че хитинът може да се намери в Chitrydiales, Monoblepharidales, Protomycetales, Hyphochyiridiales, във всички Endomycetales, Blastocladiales, Mucorales, Entomophthorales, във всички обикновени и основни гъби от тях. Оомицетите се превръщат във винят, при тези с полизахариди в присъствието на целулоза, по -често в присъствието на други гъбички.
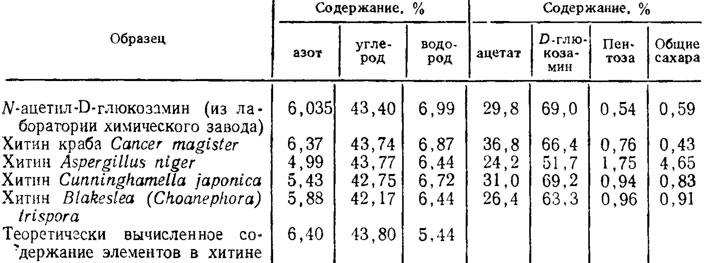
Разтягане на последния час в звука на проявите на силата практичен vicarystannyДоказано е, че генетиката на гъбичките за синтез на полимери достига много данни в по -голям мащаб за синтеза на полимери за синтеза на полимери (Таблица 1.4). Ацетатът и D-глюкозаминът са най-ниските в Aspergillus niger, тъй като се обясняват с голямото количество пентози и глюкоза в склада на β-глюкан и два α-глюкана от гъбичната обвивка. Анализ на дифракционните модели, показващи идентична кристална структура на тази на гъбички и раци с много по -голям ред в останалите. В допълнение, гъбички хитин срещу ламеларната структура на ракови хитин mav влакнести будови (Feofilova et al., 1980).
Хитозанът е заместител на хитина в някои мукозни гъбички, хитозан, който протича със слаба киселинна реакция на хидролиза до хитозанов сулфат, известен, например, Mucor rouxii, също в Phycomyces blakesleeanus. Броят на ацетиловите групи в новия беше намален до нула в M. rouxii. От другите аминополизахариди, полимери на галактозамин със здрава аминогрупа, здраво фосфатно съединение, със структура от тип хитозан, с наблюдения на клетъчни тунели Neurospora crassa, N. sitophitla, A. niger и Botrytrie 65 (A. niger и Botrytrie 65 ) Редица гъби имат полимеризация на аминозахари, асоциирани с манани, глюкани и протеини.
Целулозата в черупките на гъбите не е податлива на хитин наведнъж. Човек може да получи един Rhizidiomyces от порядъка на хипохондричните гъбички, при които вонята се появява веднага. Луковичната целулоза се среща в гъби от оомицети от порядъците Acrasiales, Lagenidiales, Saprolegniales, Leptomitales, Peronosporales (Aronson, 1965).
В средата на гъбите, които се задържат във водната среда, целулозата расте на групи, което може да причини двустранни зооспори. В Blastocladiales и Monoblepharidales, които могат да бъдат едноизгорени зооспори, тя е тъпа. Винятка, как да стана Rhizidiomyces от порядъка на Hyphochytridiales, но веднага имам хитин и целулоза и се състезавам за Nable в преходна форма между хитиновите Chytridiales и Blastocladiales цели клетки. Qia единично изгорялата форма е maê dzhgutik зооспорите са перистома, а не бич, както в Blastocladiales и Monoblepharidales. Цикаво, пъпките на перата на оперените оомицети са нагаду на бъговете на бактериите, които са подобни на тези на жгутиците.
Целулозата в гъбите се развива лесно като микрохимичен реактив на Speitzer или реагент, който може да се съхранява в количество йод в йодните калории с добавяне на 70% от количеството органична киселина.
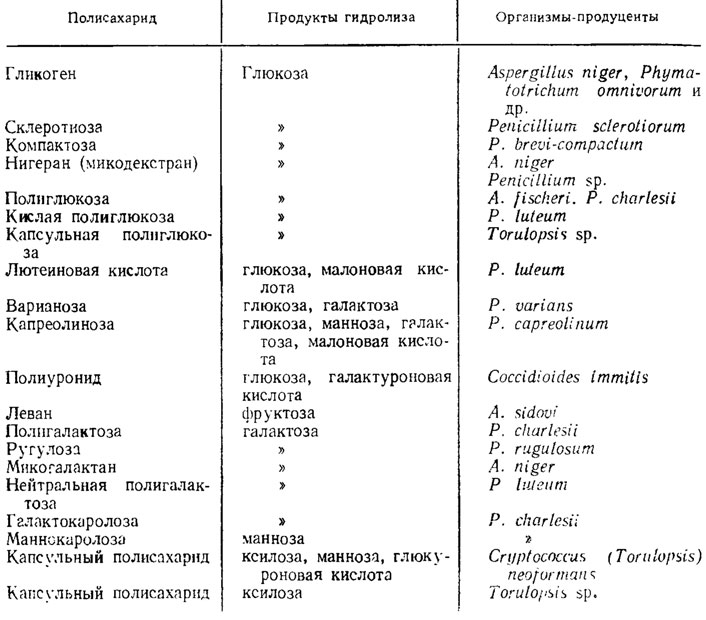
Глюканът в гъбите е още по -изразен и образува целулоза според тяхната структура. Мономерията също е глюкоза. Най -ефективните глюкани обаче са по -малко от кристалните структури, по -малко целулоза. Кристалността на Gluck-niv нараства във вкуса на установяването на хидроглюкани, когато те се обработват със сирханова киселина. Някои глюкани, които се намират във връзка с протеини, се намират в други и в Penicilliunt notatum (Aronson, 1965). Вторият глюкан, който може да влезе в склада на черупките на гъбичките, е калоза, която е подобна на появата в цитовидните тръби на растящите роси и силно разпръснати основни барбници, тоест има кисела природа, срещу целулозата е β -1,3-глюкозид. Podbny glucan, farbuvaln на основните barvniks, bauvies в Sclerofinia. Глюканът също е в Aspergillus fischeri, Allomyces macrogynus, Neurospora crassa. Гъбите също развиват хетерополни захари, съхранявани от мономери на малки бонбони, особено в рода Penicillium. В други форми, патогенни за същества, като Coccidioides и Cryptococcus, някои от тях, дори кисели полизахариди, влизат в склада на капсулите. Поставете глюкани и други продукти в полимеризацията на монозахариди и захарозни киселини, преди да влезете в магазина за клитин и резервни гъбни копия, поставете в таблицата. 1.5.
Манани, полизахариди, се съхраняват в мономери на маноси. Особено ярък е при децата, освен това вонята е част от вида деца, които се задържат на повърхността на камбиала, изправени срещу кората на дърветата. За броя на тези форми чувалът се въвежда под кората на иглолистните дървета Hansenula; В зараждащата се клетка на други полизахаридът присъства под формата на фосфоманан в капсули. Целият хидрофилен и лигавичен полизахарид се прилепва към четините наведнъж един от друг към четините, който обхваща само короядите и по този път може да се пренесе такъв път от едно дърво на друго (Wickerham and Barton, 1961). При хифеените гъби манозата не се появява, но манозата расте в хидролизати на тези клетки.
Гъбите също развиват полизахариди, разкривайки галактоза, 6-дезоксипентоза, метилпентоза, най-често фруктоза, особено лигавица. В черупките на цитин Penicillium chrysogenum са известни були от броя на пентозите на 6-дезоксипентоза, рамноза и ксилоза, а също и в гъбата Polysiictus sanguineus.
Полизахаридно-протеинови комплекси от були се появяват при деца, например при Candida albicans, причинител на млечница при кърмачета. Манан-протеинов комплекс от знания в Saccharomyces.
Гъбите са склонни да растат силно в присъствието на умове dovkillaи вику култура. Единственият обхват е 35-36% от масата на клина за суха реч. В други повече от 3% от липидите се намират в черупките на их. В черупките на мукозни гъби се открива воня, например в Mycor rouxii и Phycomyces, в спорангията на които има близо 25% липиди от сухи маси. Jmovіrno, вонята, която трябва да отмъсти в кутикулата (Aronson, 1965).
Пигментите също могат да бъдат включени в малки количества до склада на гъбните черупки. Пигментите, особено често локализирани във виглиадите на специална топка, черен пигмент, меланин, се намират още по-често в клетките на мицела или в черупките на супер-потока от гъби багатох. Такава меланинова топка е в черупките на аскоспорите на Neurospora tetrasperma (Aronson, 1965).
Меланинът се наблюдава при гъби с ниска активност на полифенолоксидаза, тъй като участва в тази биосинтеза, както и в обмена на активни дехидрогенази. Такива гъби, включително представители на родовете Fusarium, Trichothecium, Arthrobotris, Cephalosporium и много антибиотици, се характеризират с най -честите еризипели и портокали в спороносните структури, лежат в ролята на голямо количество минерали ... Връзката на проявата на каротеноиди виски ривнемАктивността на дехидрогеназите се обяснява с факта, че оптималният окислително-основен режим за дехидрогенази се използва за мозъка, който е оптимален за биосинтеза на каротеноиди и други продукти на шунта на терпеноидите. Каротичните пигменти не влизат в хранилището на клетъчните мембрани на гъбичките, а се локализират или в клетъчните мембрани, или в петна от липиди, зачервени в розово в цитоплазмата. В близост до гъбички се наблюдават абсолютно специални пигменти, като оммохромия, много пигментни коми, в гъбата Pycnoporus (Polyporus) cinnabarinus (Shivrina, 1965) или системата на фикобилини от вида сироаденоиди Russ. Фотосинтез в синьо- зелени и червени водорасли.
МИКРОБИОЛОГИЯ 2010, том 79, бр.6, стр. 723-733
УДК 582.281 (047)
КЛИТИННА СТЕНА ОТ ГЪБИ: СУХАСНИ ОБЯВИ ЗА СКЛАДОВЕ І БИОЛОГИЧНИ ФУНКЦИИ
© 2010 Е. П. Феофилова
настройвам Руска академияНаук Институт по микробиология ИМ. S.N. Виноградски РАН, Москва
Надежда до редактора 05.11.2009 г. r
Инспекция на заданията на малки vyvchennoy повърхностни структури на клетки от мицелни гъбички - клетъчна линия (CS). Водени от данните за методите за наблюдение и преобразуване на чистотата във видимостта на цитоплазмата вместо на фракцията KS и за химическия склад. Докладвани са структурни (скелетни) и вътрешни структурни компоненти на CS- аминополисахарид, а- и v-глюкан, млечен, липид, уронова киселина, хидрофобен, спорополенин и меланин. Особено уважение има хитинът, новата му функция в антистресовото лечение на клетките, както и индикациите за някои аминополисахаридни гъби в хитин от водорасли и членестоноги. Обсъжда се явлението апикален растеж на хифите и съдбата на специални микровезикули в морфогенезата гъби клитини... Воден от познанията за ензимите, той ще участва в синтеза и лизиса на CS. В крайна сметка се обсъжда функционалното значение на KS в гъбите в контекста на повърхностните структури на гъбата.
Ключови думи: микроклетъчни гъби, винена клетка, методи за визуализация, химически склад, апикален растеж, физиологични функции, морфогенеза.
Един от ключовите проблеми съвременна биологияе Хранене - като ранг в процеса на развитие, установяването на морфологично същите организми, като биохимични механизми и като клинични структури, участват в цял процес, който е срещу планетарната скала. Не по -малко са данните за тези, които вливат стрес върху морфогенезата и биополимерите, за да контролират формата на клитина. Як показват резултатите и doslіdzhen останалата част от скалата, Нарастващата хифа на микроклетъчните гъбички е уникален модел, който позволява визуализация на процеса на образуване на клетка и това, тъй като запазва образуването на клетъчна морфология в онтогенеза. Друго голямо допълнение към предварителното развитие на явлението са направили данните за еволюцията на склада и биологичната функция на CC на мицелиалните гъби.
KS roslin куршум открита през 1665 rotsі, и KS гъби само на кочана 18 -ти век. Въпреки това, повърхностната структура на тривиален час на практика не се вписва, така че беше важно CS viconuê само да "рамкира" функцията и те ощипиха кабината със стените, които трябва да поддържат опората. В началото на 20 -ти век позицията преди COP се променяше бързо и щеше да бъде много бавна, но най -вече в случай на отглеждане на растения и бактерии. Интензивното развитие на гъбичките и биотехнологичните виробници, при които производителите на биологично активни вещества измиват мицелни гъби, засилва розата
1 Адреси за кореспонденция (имейл: [Имейл защитен]).
ча научен дослиженот vivchennyu KS гъби. Данни за повърхностните структури за периода до 80 -те години на миналия век, побойникът узагални в първия в светлината на книгата за KS гъбите. В офанзивната скала основният набор от роботи е паднал до края на ХХ и началото на XXI век. Bully извърши основно предварително третиране, възлагане на систематичност, химическо съхранение на KS, апикален растеж на хифити, метаболизъм на хитин, антихигроскопични лекарства, хидрофоби, ковалентно свързваща кръвна захар, ензими, витамини Въпреки че не се говори, но не се обсъжда в крайна сметка, е възможно да се покаже, че повърхностната структура на клиента не е многофункционална и не е многофункционална, поради ролята на процеса, който не е отговорен за болестта. и адхезивна сила, контрол на процесите на диморфизъм и обхваща растежа на гъбични клетки, освобождаване на сигналния сигнал и прехвърлянето му към мембраната и вътрешните пратеници. Ще погледна назад към казаното, което уважавахме за приключването на дискусията за предстоящите дати: за склада на химията на CC, биологичните функции на основните биополимери, бъдещето на CC и процесите на ферментация с много растежи в хифите на гъбите, COP.
Химически склад КЛИТИНО
ФУНКЦИЯ СТИНОК ГРИБИВ ТА БИОЛОГИЧНА ЇЇ ОСНОВНИ КОМПОНЕНТИ
Възстановяването на химически склад на KS гъби се ремонтира от отстраняването на чиста част от структурата на структурата, за да се пречисти KS като цитоплазмена обструкция. Първата процедура е премахването на гъбичките и образуването на гъбички на първо място - да се работи при ниски температури (близо до 4 5 ° C), за да се предотврати влошаването на разграждащите ензими. Преди ruinuvannyam, виното се замразява при температура близка до азота. Ruynuvannya klіtin zdіysnyuêtsya или в специални хомогенизатори, или на преси по метода "твърд порок". Добрите резултати са още по -далеч от ултразвуковия дезинтегратор. Визуализацията на студената вода вместо на студената се повтаря 4-5 пъти. За по -голяма визуализация на цитоплазмата на мястото на вискозата, видимостта на викории No C1, 8 М сечовини, 1 М амоняк или 0,5 М оцетна киселина. В настоящия час методите за визуализиране на гъбичното CW в процеса на онтогенеза бяха разбити и освен това методите бързо се развиват за микроцелия и постелката. По -късно отидете директно на дозировката, zokrem, с наличието на полизахаридни съхранение, от KS vityagayut lípіdi, vikoristovuyu органични рози, например, хлороформ и метанол в spívіdnіtníy 2: 1, добър резултат, Tsei razchinnik добро pidsushuê KS и по един начин може да замести лиофилното сушене. Офанзивният етап е стойността на чистотата на отряманата фракция на КС. Използвам техниката на vikoristovuyu svitlov и електронна микроскопия и специални методи за подготовка. Найбилш често застоява реакцията въз основа на връзката I3- с хитозан. Изолираната чиста KS с решението на Lugol ще бъде заредена в рима или виолетов цвят, в този час, когато непокътнатата KS може да бъде заредена. Можете също така да бъдете тествани за наличие на DAPI ядра (4,6-yokt1ёto-2-rkepuTsn-yoo1). Особено важно е да се разглежда чистата фракция на КС като обща процедура, за да се определи началото на резултатите и анализът на химическия склад. Като цяло, в случай на представител на мукозни гъби, в мицелата на робота е открит глюкан, по-точно комплекс от хитозан-глюкан, бих искал всички предварително да знаят за наличието на муколи в мицела на глюкан.
За даден час компонентите на гъбичния CW се добавят към структурните компоненти (хитин, р- (13) -р (1-6) -глюкани, р- (1-4) -глюкан (целулоза) и вътрешни структурни (те наричат матрицата), към които стоманени форми на манопротеин, галакто-манопротеин, ксилит-манопротеин, глюкурон
манопротеин и а- (1-3) -глюкан. Nimensh vivcheni a-glucan, който е линеен полимер на глюкоза (в Schizosaccharomyces pombe, такъв глюкан може да бъде отстранен от около 260 излишни глюкози). Целият полимер се съхранява в две плетени ленени копия, така че близо 120 (1-3) -a ^ -глюкоза излишъци и (1-4) -a-D-глюкоза излишъци по краищата на полимерната молекула. Признавайки, че глюканът е необходим за осигуряване на процеса на морфогенеза. От CS мицела на Penicillium roqueforti във водата се забелязва присъщ глюкан, тъй като той виси на ливадата, но се вижда разлика от 10% разлика в количеството литиев хлорид в Dimox-tidel. Когато глюкан се инжектира по метода на метилиране, булите се елиминират две глюкоза, получена от три-О-метил, които показват връзките 1-3 и 1-"Този резултат е показателен за линейната структура на молекулите, така че само Показана е връзка 1 - " - 3 и 1 -" - 4 с излишък на глюкоза. Данните бяха потвърдени чрез анализа на 13С-ЯМР спектъра на полизахарида, който също доведе до конфигурацията на цялата излишна глюкоза. Такива полизахариди, особено за химическа модификация (сулфатиране и карбозиметрична баня), с добро качество във вода, могат да бъдат полезни като биологично активни полимери и да станат интересни за витамините
По -ниски във вода (1 -" -3) -a ^ -глюканови куршуми бяха забелязани по -рано от редица видове ядливи и по -ниски гъби, включително Penicillium chrysoge -num, и местни a ^ -глюкани от два типа връзки (1 - 3) і (1- "- 4), който расте в съпрузите, са били известни в клиновете (Aconi-tum kusnezoffii Reichb).
Структурният компонент - целулозата - е характерен за оомицетните гъби, които според сегашната таксономия се довеждат до псевдо грозде. Складът на монозахаридите на KS включва глюкоза, маноза, ксилоза и освен това прехвърляемата захар е глюкоза, като склад до 68%. Аминокиселини, липиди (не повече от 3%) и ацетил-глюкозамин също трябва да бъдат включени в склада на гъбите KS. Глюкан, например 1,3-глюкан, се използва за образуване на микрокристален комплекс с хитин, заглавието на хитин-глюкановия комплекс (CHC), който може да се превърне в гъба. Целият комплекс от присъствие в COP е практичен за всички гъби, с вината на зигомицетите, да служи като високо систематично запознаване. При някои гъби по повърхността на CS се открива лигавичен материал, който може да се съхранява в полизахариди, изображения на ß-1,3-глюкани, които могат да заменят само глюкоза от един тип, с ß-1,6-връзки от дермалната трета глюкоза на един
Склад за химияи структурата на клетъчната линия на гъбичките
Клас гъби багатошаров, с голямо разнообразие от одобрени версии, се разпространява в склад по химия със структурни въглехидрати, като склад по химия може да бъде разпределен в 3 групи:
полимерна глюкоза(Глюкан, хитин, целулоза). Glucan ще съхранява голяма топка от големите гъби. Вътрешната топка на канцеларския стил на гъбите на lantsyuzhkami претендира за хитин, natayuzhi zhorstkist. Хитинът замества целулозата, Як в голям брой гъби всеки ден или дори влиза в склада на оомицети, тъй като за даден час няма типични гъби. Деацетилиран хитин ще го нарека хитозан, който в комплекса с хитин определя клетъчната линия на зигомицетите.
полимерни монозахариди (маноси, галактоза и др.)на vidmіnu от vishchikh roslin, de voni формират основата на матрицата píd по имехемицелулоза, по -малко типична за гъбичките. Винятката става дирижи, в клитиновите уили, които са особено богати на полимери на маноси, наречени манани. Pripuskayut, такъв склад на стената ще бъде по -красив от brunkuvannya.
полимери на въглехидрати, ковалентно свързани с пептиди (гликопротеин)да оформя средната топка на голяма топка култура и да играе важна роля, както в развитието на структурната цялост на клетките, така и в комуникационните процеси с новата среда.
Към следните специфични характеристики се прилагат гъбни клетки: видимост на пластидите, Shcho zblizhuê я с клин от същества;
видимостта на нишестетоЗа Eumicetes той се заменя с полизахарид, близък до нишестения гликоген на създанието, при оомицетите - с полизахарид, близък до Laminarin Buri. Извършват се редица въглехидрати за съхранение, специфични за гъбичките.
viroblenya на специфични вторични метаболитиРолята на антибиотиците, фито- и микотоксините, фитохормоните е по-важна.
Той се прилага и за специфични характеристики на гъбите хетерокариаза и парасексуален процес.
При гъбите проявата на хетерокариаза е дори по -широка или на различни ядра, като ядрата се събират в една клетка за тривиален час, хетероаллелност за някои гени. Хетерокариазата замества хетерозиготността и адаптациите на гъбите към младите умове за хаплоидни гъби. Доказателствата за разнообразието от ядра са обобщени от редица специфични характеристики на гъбите:
1. наличието на повече от едно ядро в клетка
2. специфична Будова на клитиновите прегради, в един или повече малки отвори, наречени от порите, през ядрата можете да преминете от един клитин към theinshu
3. Gifs в средата на същите колонии и за намиране на млади, близко израснали колонии, които са израснали от възрастното супер оръжие на един вид гъби, често растат, в резултат на което те могат да обменят ядра на по-стари щамове.
Парасексуален (псевдосексуален)процес. Дори в хетерокаротични клетки ядрата са гневни, хетероаллелност от определен локус и хетерозиготно диплоидно ядро. Можете да ядете в превъзходно и ухо от дата на диплоиден хетерозиготен клон. В процеса на митоза диплоидните ядра могат да се превърнат в хаплоиден лагер, например, в един набор от хромозоми, в противен случай те могат да се обменят с хромозоми (митотично пресичане). Нападателните процеси се контролират чрез рекомбинация на гените и фенотипите на Баткив. Парассексуалната (без скоби) рекомбинация е още по-рядка, тъй като не изпреварвам едно ядро на милион, дори ако има голям брой ядра в мицела, тя постоянно се насърчава в популации от гъби.
развъждане- вегетативно, без гражданство, щатски.
вегетативно- фрагментация на талуса, осветяване на хламидоспори, тъй като те растат спокойно в мицела, като наред с други образуват синини.
без гражданствоРазмножаването в млади гъби може да бъде създадено чрез разпадане и непокорни спори. Зооспорите допускат относително малка група гъбички и гъбоподобни организми - вода и земя, в които генетичните връзки с водни гъби и водорасли са ясно видими. Структурата на jgutik е важно диагностично познание по време на въвеждането на царството на предпеенето. В случай на голям брой гъби, с разпространение без гражданство, има жалък спор за това, което се е случило със земята от дълго време. За момент се развива формирането и локализацията на ендогенното развитие, установява се в спорангии спорангиоспори и екзогенни (конидии), които се развиват върху специални хифи - конидиофори. Конидиите се установяват в големи гъби (Ascomyceti, Basidiomyceti, Deyaki Zygomyceti), съхранение на конидиално спорообразуване, което води до по -голям растеж и широко развитие на гъбите.
Състояние на размножаване на гъбиИмам своя собствена специфика, както в морфологията на законоустановения процес, така и в механизмите на генетично и физиологично регулиране на статусите и предаване на информация за разпадане.
соматогамия- най -разпространеният и най -простият тип статистически процес, полето в zlitty не се диференцира в гамети на соматични клетки. Inodí vin противікê да навит без яд китин - ядрата в средата китини се ядосват. Израствайте при великите базидиомицети, общи други и дела. Таксонов.
гаметангиогамия- гаметангиите се появяват върху хаплоидната мицелия; Такъв законоустановен процес е характерен за повечето обикновени гъби. Вариант на гаметангиогамията е зигогамията при зигомицетите.
гаметогамияпри viglyad z-hetero и oogamy гъбите се развиват значително повече, по-ниско при другите евкароти. Изо- и хетерогамията се развива в хитридиомицетите. Класическата оогамия с изявленията на сперматозоидите и ооцитите в гъбите не се върти, но има силна промяна в опциите.
Поради особеностите на регулирането на статиката в гъбите съществуват редица видове статистически процеси
ГинандромиксисМоже да се види на дупето на двуводни оомицети, в някои оогонии и антеридии те се развиват върху малки слоеве, например фитофлуор или картофолова гъба. Ако това е генетично едностранно щам на viroshuvati в монокултурата, тогава той ще се размножава само по начин без гражданство. Веднага след като бъдат инструктирани да опишат мецелия на два щама, тогава в тях ще се появят морфогенетични змии от другите химически видения със стероидна природа - законоустановени феромони. Anteridol иnduku одобрение на антеридия от партньор, и oogonіol - oogonіev. В същото време регулирането на статиса е от голямо значение: ако щамовете са в състояние да установят антеридии или оогонии, да депозират някои от най -често срещаните феромони при нов и ти партньор. Звуци Името на процеса на изготвяне е ginandromixis.
Димиксис или хетеротализъм.Отдавна е отбелязано, че гъбите могат да бъдат хомо-чи хетеротални. При хомотални видове по време на статичния процес генетично идентичните ядра на всички средни мицели се ядосват. При хетеротални видове за преминаването на статистическия цикъл е необходимо за различен етап (за a нови видовегъби) ядосани върху областите на супер-потока (по-точно техните ядра). За душевното състояние на двата щама е необходимо генетично да се разминават (хетероалалност) в пеещите локуси, наречени локуси за сдвояване. Повечето гъби (зигомицети, аскомицети, част от базидиомицети) имат едно място на чифтосване с два алела. Локусът на сдвояване се основава на децилни гени, които се основават на синтеза на задължителни феромони. Такъв хетеротализъм се нарича еднофакторен или биполярен. Потомството на такива гъби, за мейоза, прераства в две несъответствия, макар и взаимно консумируеми групи в съотношение 1: 1, така че местните (инбридинг) и неразмножаващите се
В геномите на годни за консумация базидиомицети има два локуса на сдвояване - A и B, а сумата на един от единствените е хетероаллелна и в двата локуса (Ax Bx сумира от Ay By, но не от Axe By и Ay Bx). Такъв хетеротализъм се нарича двуфакторен или тетраполярен. Процентът на печалба при инбридинг се намалява с до 25%.
Диафоромиксис- при обикновените базидиомицети не са два, а по -скоро изобилни алели към локуса на сдвояването, изпадащи в изстричане в младите щамове, така че те се натрупват в популацията. Такъв контрол на сдвояването ще осигури 100% от степента на аутбридинг, тъй като това става с помощта на различни алели, които са взаимно съвместими, а алелите са още по -изобилни. Резултатът е хибридни популации на panmix.
Жизнен цикъл на гъбитекакто и гъби и гъби. Основни цикли, аксесоар към гъбите
1. Безболезнен цикъл, характерен за величествена група от непонятни хора, които са загубили статута на репродукция. Ядрата на Розподил обикновено са митотични. Важно е, че голям брой гъби се въвеждат в най -често срещаните гъби или заедно с втория статистически процес, за да образуват формална група от непълни гъби или дейтеромицети.
2. Хаплоиден цикъл. Вегетативни са метали в нехаплоидни ядра. За държавния процес (сингамия) диплоидната зигота (охлаждане през периода на затишие) продължава мейотично - зиготична мейоза. Характерно за зигомицети и багатиохитритриомицети.
3. Хаплоидно-дикаротичният цикъл се характеризира с времето, когато изпраща зло вместо гаметангиите (гаметангиогамията) или соматичните клетки на хаплоидния мицел (соматогамия) на ядрото приемат дикариони (париетални) генетично. Вонята протича в синхрон, създавайки дикариотизираща мицела. Държавният процес се завършва от ядрените дикарионни ядра, така че зиготата се преструва, че продължава с мейоза без период на спокойствие. Мейоспорите съхраняват състоянието на образуване на спори на обикновени и базидиални гъби във вигляди аскоспори и базидиоспори. Когато те покълнат, се осъществява хаплоидният мецелий. В голям брой обикновени гъби (сред другите гъбички и гъби тафрин) в жизнения цикъл, хаплоидната фаза във виглиади вегетативния мицел (Anamorph), каротидната фаза на краткосрочните хипогастрални гъби е представена Базидиалните гъби имат диаротична фаза в жизнения цикъл, хаплоидната фаза е краткотрайна.
4. хаплоидно-диплоидният цикъл в изоморфните изкривявания на гъбите расте все повече и повече (деяки диджиджи и водни хитридиомицети).
5. Диплоиден цикъл, характерен за оомицетите и някои често срещани други. Вегетативни са диплоиден метал, гаметична мейоза, оцветяване под формата на гаметангии или гамети.
3. екологични групигъби.
Гъбите и гъбоподобните организми навлизат в земните и водните екосистеми, тъй като новата част от хетеротрофния блок, заедно с бактериите, заемат трофичното ниво на редуктори. Широкото разширяване на гъбите в биосферата се основава на редица най -важни характеристики:
1. Наличието на голяма микроклетъчна структура на талуса. (Позволява ви бързо да хванете субстрата, майка до страхотна повърхност, контакт със средата).
2. Голяма бързина на растеж и размножаване, която позволява за кратък период от време да се засели голяма маса субстрат, което води до голям брой спорове и ги разширява в големи времена.
3. Метаболитна активност на Висока, която се проявява в широк диапазон от стойности на екологични фактори.
4. Видимостта на генетичната рекомбинация означава биохимия, екологична пластичност.
5. Zdatn_st shvidko отидете в състояние на анабиоза, преживейте неприятния ум и се разтегнете за тривиален час.
клошар екологичен факторза гъби е харчов субстрат. Според съответния фактор те виждат основните групи гъби, които се наричат трофични групи.
4. къса характеристика viddіlіv гъби.