Скорочувальна вакуоль. Скорочувальна вакуоль і її фукция
Дана стаття ознайомить читача з будовою найпростіших організмів, а саме - акцентує увагу на будову скорочувальної вакуолі, яка виконує видільну (і не тільки) функцію, розповість про значення найпростіших і опише способи їх існування в навколишньому середовищі.
Скорочувальна вакуоль. поняття
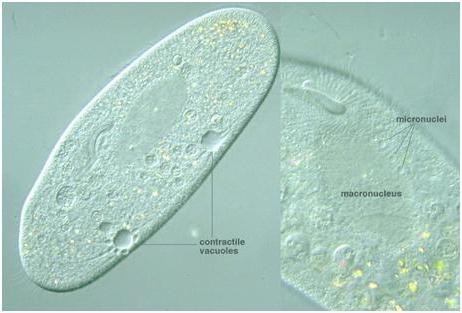
Вакуоль (від франц. Vacuole, від латинського слова vacuus - порожній), кулястої форми невеликі порожнини в рослинних і тваринних клітинах або одноклітинних організмах. Скоротливі вакуолі в першу чергу поширені серед найпростіших організмів, які мешкають в прісноводної воді, наприклад, серед протистов, таких як амеба протей і інфузорія туфелька, яка отримала таку оригінальну назву через форми тіла, схожого з формою підошви туфлі. Крім перерахованих найпростіших, ідентичні структури також були виявлені і в клітинах різних прісноводних губок, які належать до сімейства бодягові.
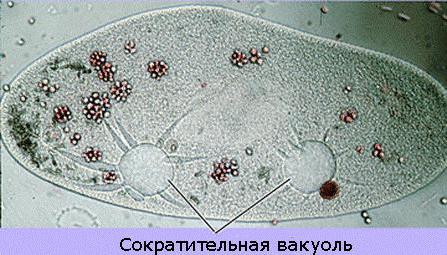
Будова скорочувальної вакуолі. її особливості
скорочувальна вакуольє мембранним органоїдом, який здійснює викид зайвої рідини з цитоплазми. Локалізація і будова цього апарату варіюється у різних мікроорганізмів. З комплексу бульбашковидних або трубчастих вакуолей, званих Спонг, рідина потрапляє в скоротливу вакуоль. Завдяки постійній роботі цієї системи підтримується стабільний обсяг клітини. У найпростіших є скоротливі вакуолі, які представляють собою апарат, який регулює осмотичний тиск, а також службовець для виділення з організму продуктів розпаду. Тіло найпростіших складається всього лише з однієї клітини, яка, в свою чергу, здійснює всі необхідні життєві функції. Представники цього подцарства, такі як інфузорія туфелька, амеба звичайна, інші одноклітинні мають всі властивості самостійного організму.
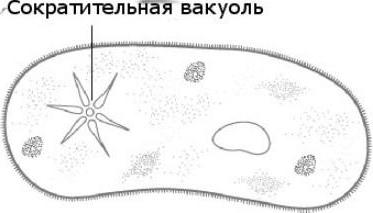
Роль найпростіших організмів
Клітка виконує всі життєві функції: виділення, дихання, подразливість, рух, розмноження, обмін речовин. Найпростіші поширені повсюдно. Найбільша кількість видів мешкає в морських і прісних водах, багато населяють вологий грунт, Можуть вражати рослини, жити в тілах багатоклітинних тварин і людини. У природі найпростіші виконують санітарну роль, також вони беруть участь в круговороті речовин, є їжею для багатьох тварин.
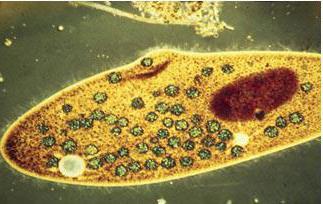
Скорочувальна вакуоль у амеби звичайної
Амеба звичайна - представник класу корненожки, не має на відміну від інших представників постійної форми тіла. Пересування здійснює за допомогою ложноножек. Тепер розберемося з тим, яку функцію виконує сократительная вакуоль у амеби. Це регуляція рівня осмотичного тиску всередині її клітини. Вона у може утворитися в будь-якій ділянці клітини. Через зовнішню мембрану вода з навколишнього середовища надходить всередину осмотически. Концентрація розчинених речовин в клітині амеби вище, ніж у навколишньому середовищі. Таким чином, створюється різниця тиску всередині клітини найпростішого і за її межами. Функції скорочувальної вакуолі у амеби - це своєрідний відкачує апарат, який виводить надлишок води з клітини найпростішого організму. Викидати в навколишнє середовище накопичилася рідина амеба протей може в будь-якій ділянці поверхні тіла.
Крім осморегуляторной, виконує функцію дихання в життєдіяльності, так як в результаті осмосу надходить вода доставляє розчинений в ній кисень. Яку ж ще функцію виконує сократительная вакуоль? Так само виконує функцію виділення, а саме разом з водою виводяться продукти обміну речовин в навколишнє їх середовище.
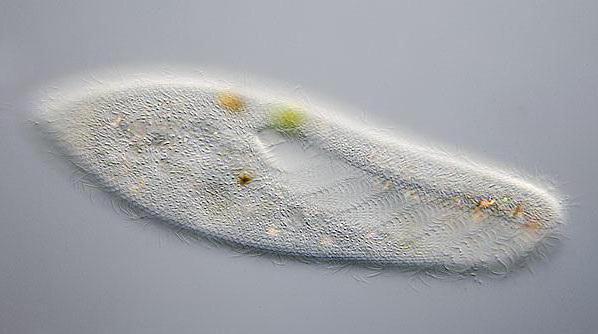
Дихання, виділення, осморегуляція у інфузорії туфельки
Тіло найпростіших покрито щільною оболонкою, яка має постійну форму. так і водоростями, в тому числі і деякими найпростішими. Організм інфузорії має більш складне, ніж у амеби будова. У клітці туфельки спереду і ззаду розташовані дві скоротливі вакуолі. У цьому апараті помітні резервуар і кілька невеликих канальців. Скоротливі вакуолі постійно знаходяться, завдяки такій будові (з мікротрубочок), на постійному місці в клітці.
Головна функція скоротливої вакуолі в життєдіяльності даного представника найпростіших - осморегуляція, також вона виводить з клітки зайву воду, яка проникає в клітину за рахунок осмосу. Спочатку відбувається набухання каналів, що приводять, потім вода з них перекачується в спеціальний резервуар. Резервуар скорочується, відділяється від каналів, що приводять, через пори вода викидається назовні. У клітці інфузорії знаходиться дві скоротливі вакуолі, які, в свою чергу, діють в протифазі. За рахунок роботи двох таких апаратів забезпечується безперервний процес. Крім цього, вода безперервно циркулює завдяки діяльності скорочувальних вакуолей. Вони стискаються по черзі, і частота скорочень залежить від температури навколишнього середовища.
Так, при кімнатній температурі(+18 - +20 градусів за Цельсієм) частота скорочень вакуолей становить, за деякими даними, 10-15 секунд. А враховуючи, що природним місцем існування туфельки є будь-які прісні водойми зі стоячою водою та наявністю в ній розкладаються органічних речовин, Температура цього середовища на кілька градусів змінюється в залежності від пори року і, отже, частота скорочень може досягати 20-25 секунд. За годину сократительная вакуоль найпростішого організму здатна викинути з клітки воду в кількості. порівнянній з її розмірами. У них накопичуються поживні речовини, неперетравлені залишки їжі, кінцеві продукти обміну речовин, також можна знайти кисень і азот.
Очищення стічних вод найпростішими
Вплив найпростіших на круговорот речовин в природі має величезне значення. У водоймах, внаслідок спуску стічних вод, Розмножуються у великій кількості бактерії. В результаті з'являються різні найпростіші організми, які і використовують в їжу ці бактерії і таким чином сприяють природної
висновок
Незважаючи на просту будову цих одноклітинних організмів, тіло яких складається з однієї клітини, але виконує функції цілого організму, дивним чином пристосованого до навколишнього середовища. Це можна спостерігати навіть на прикладі будови скорочувальної вакуолі. На сьогоднішній день вже доведено величезне значення найпростіших у природі та участь їх у кругообігу речовин.
А. скорочувальна вакуоля
1. Морфологія скорочувальних вакуолей
Більшість найпростіших (крім Sporozoa)мають особливий, більш-менш строго локалізованим, іноді вельми складно влаштованим екскреторне апаратом, здебільшого мають характер так званих скорочувальних, або пульсуючих, вакуолей.
як загальне правило, Вакуолі є у всіх прісноводних Mastigophoraі Sarcodinaі у всіх інфузорій.
У Mastigophoraздебільшого є всього 1 сократительная вакуоль, рідко 2 (Phytomastigina).
Розміри вакуолярного апарату в загальному прямо пропорційні розмірам тіла і обернено пропорційні кількості вакуолей в тілі тварини. Так, у великих інфузорій Heterotricha (Stentor, Spirostomum)є всього 1 вакуоль, але зате вона (разом з її каналами) досягає дуже великих розмірів. У Trachelius ovumдо 30 вакуолей, але зате вони дуже дрібні.
Іноді помічається збільшення числа вакуолей в міру росту тварини, як наприклад у Collinia branchiarumі деяких інших Astomata.Те множення числа вакуолей, яке у Collinia (Anoplophrya)відбувається протягом індивідуального життя, мало, мабуть, місце протягом еволюції всієї групи Astomata(Рис. 124). Дійсно, серед них можна намітити ряд форм з поступово ускладнюються вакуолярного апаратом. Так, у Dogielellaє всього 1 субтермінально розташована вакуоль, у роду Anoplophrya 1 поздовжній ряд з невеликого числа вакуолей; у пологів Butschliellaі Мопоdontophryaчисло вакуолей в цьому ряду сильно збільшується, а у Radiophryaвакуолі утворюють 2 поздовжніх ряди. Нарешті, у роду Haptophryaвідбувається злиття ряду численних вакуолей в загальний поздовжній вакуолярного канал, спосіб походження якого видно з наявності уздовж нього цілого ряду окремих вивідних отворів.
У Sarcodinaі Mastigophoraвесь екскреторної апарат обмежується самою вакуолью. У небагатьох жгутіконосцев (наприклад, Euglena)скорочувальна вакуоль оточена віночком дрібних освітніх вакуолей, з'єднаних з пий тонкими канальцями.
Паппас і Бранд (Pappas a. Brandt, 1958) думають, що вода в вакуоль у Amoebaнадходить внаслідок виливу в неї дрібних бульбашок, що лежать навколо, шляхом злиття мембран цих бульбашок з центральної вакуолью. У інфузорій зазвичай скоротливі вакуолі забезпечуються одним або декількома приводять каналами, які пульсують. У Spirostomum - 1 такий канал, у Stentor- 2, у Frontonia- IO довгих каналів і т. Д. Канали, по-видимому, розходяться в цитоплазмі на набагато більш далеку відстань, ніж це видно під мікроскопом. По крайней мере, при осмірованіі Parameciumстає помітним продовження периферичних кінців каналів в довгі, тонкі концевиe відділи. Освітні вакуолі і призводять канали збирають рідина (т. Е. Головним чином воду) з цитоплазми і доставляють її в скоротливу вакуоль. Весь вакуолярного апарат інфузорій залягає в глибині ендоплазми і не змінює свого положення в тілі.
Нещодавно Шнейдер (Schneider, 1960) опублікував цікаві дані по ультрамікроскопічних будовою скорочувальної вакуолі і каналів, що приводять Paramecium caudatum.За даними Д. H. Насонова (1924), що призводять канали оточені осміофільние муфтою, а по
спостереженнями Гелея (Gelei, 1925а, 1925b), губчастої плазмою (Nierenplasma або Nephridialplasma).
За дослідженнями Шнейдера, ця ділянка плазми, що оточує призводять канали, складається з численних тонких і звивистих
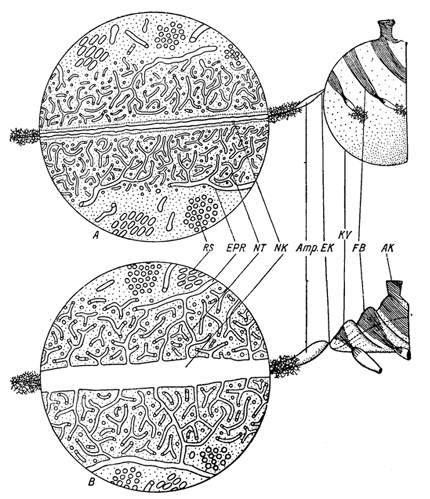
Мал. 125. Схема будови скорочувальної вакуолі з призводить каналом і навколишнього його цитоплазмою у Paramecium за даними електронної мікроскопії. (За Schneider, 1960). А - радіальний канал в стадії систоли, скорочувальна вакуоль в стадії діастоли; В - радіальний канал в діастолі, скорочувальна вакуоль в систоле. AK - вивідний канал вакуолі; Amp - ампула периферичного каналу; EK - канал, який з'єднує ампулу з скорочувальної вакуолью; EPR - канали ендоплазматичноїмережі; FB - скоротливі фібрили вакуолі; KV - скорочувальна вакуоль; NK -пріводящій канал скорочувальної вакуолі; NT - мережа "нефрідіальних каналів", пов'язана з каналамиендоплазматичної мережі; RS - скупчення "нефрідіальних каналів" в поперечному розрізі.
трубочок з осміофільние стінками (рис. 125). Внутрішній просвіт цих трубочок під час систоли дорівнює 100 Å, а під час діастоли 150 Å. Весь шар нефрідіальной плазми має товщину 1-1,5 μ. На його периферії трубочки переходять безпосередньо в канальці ендоплазматичної мережі, які пронизують все тіло парамеции. Крім того, нефрідіальная плазма оточена ще особливими трубочками з осміофільние стінками. Вони розташовуються іноді пучками. Їх діаметр досягає 500 Å. Довжина цих трубочок близько 1 μ.
Приводить канал має осміофільние оболонку. Під час систоли цей канал стає щілинним і має просвіт близько 300 Å, в діастолі він розширюється до 4000 Å. Трубочки, що оточують приводить канал, відкриваються в нього тільки під час діастоли, коли вони збільшуються в діаметрі, тоді як під час систоли ці трубочки замкнуті і не пов'язані з каналом. Приводить канал на своєму проксимальному кінці безпосередньо переходить в ампулу, яка через кінцевий каналец відкривається в резервуар вакуолі. У стінці ампули і кінцевого каналу залягають трубковидную скоротні фібрили, розташовані кількома пучками. З стінок кінцевого каналу вони переходять безпосередньо на стінку резервуара скорочувальної вакуолі, розподіляючись там також у вигляді окремих пучків.
За даними Руджінской (Rudzinska, 1957), в цитоплазмі навколо вакуолі Tokophrya infusionumзосереджуються дрібні бульбашки ергастоплазма і скупчуються мітохондрії. Крім того, навколо вакуолі виявляються структури, які можна порівняти з діктіосоми. Вони, мабуть, беруть участь в секреції і накопиченні рідини, що надходить з цитоплазми.
А. А. Стрільців (1939) знайшов навколо вивідного каналу у Cycloposthiumдобре помітне кільцеве волокно, що представляє собою, по всій ймовірності, Міон, регулюючу вихід рідини через канал назовні. У всіх інфузорій екскреторну отвір скорочувальної вакуолі є Преформовані пору. Зазвичай воно округле або овальне і має абсолютно постійне для кожного виду положення серед рядів війок. У роду Conchophthirusотвір має вигляд вузької щілини з потовщеним краєм. Бути може, така форма пори у Conchophthirusмає відоме функціональне значення при її закриванні. А саме, досить імовірно, що тут закривання відбувається не за допомогою кільцевої перетяжки вивідного каналу, а спадением країв щілини на зразок двох губ. Пора відкривається в вивідний канал, зазвичай дуже короткий, але іноді (у Paramecium trichium)утворює кілька петлевидних вигинів.
Електронномікроскопіческое дослідження скорочувальної вакуолі Tokophrya infusionumпідтвердило наявність постійно існуючої пори і екськреторного каналу, навколо якого розташовуються фібрили (180 А товщиною), які здійснюють його стиснення (Rudzinska, 1957). Аналогічні фібрили виявлені у Paramecium(Schneider, 1960).
2. Функціональне значення скорочувальних вакуолей
1) Процес скорочення пульсуючих вакуолей
Вакуолі працюють ритмічно, поперемінно то розширюючись (стадія діастоли) і повільно наповнюючись рідиною, то раптово скорочуючись (стадія систоли) і виштовхуючи свій вміст через вивідний канал назовні. Там, де є особливі призводять канали, цикл скорочення всього апарату ускладнюється; а саме, він починається з розширення каналів, що відбувається в той час, коли вакуоль перебуває в стані систоли (діастола каналів); за цим слід скорочення каналів (систоли), виливають свій вміст в скоротливу вакуоль, що викликає її систолу; в той же час канали починають знову розширюватися. Фактично відповідні стадії є і у вакуолей, позбавлених системи каналів, але там вони мало помітні або зовсім не видно, так як при діастолі вакуолі в неї виливаються знаходяться в навколишньому плазмі дуже дрібні і неправильно розташовані вакуолькі (пор. Ophryoscolecidae,Мал. 126).
Темп пульсації варіює у різних найпростіших, а також змінюється і у одного і того ж виду під впливом зміни факторів зовнішнього середовища.
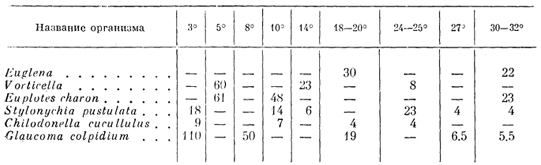
Так, для різних прісноводних Protozoaпроміжки між пульсаціями при різних температурах показані в табл. 6.
ТАБЛИЦЯ 6
Зміна темпу пульсації скорочувальної вакуолі у інфузорій(В сек.) при різних температурах
Oligochaeta.При цьому виявилося, що темп пульсації скорочувальної вакуолі не є однаковим у всіх Astomataі знаходиться у зв'язку з осмотическими умовами того середовища, в якій мешкає господар. У Mesnilella clavataw. Hoplitophrya secansз прісноводних олигохет пульсація настільки ж часта, як і у вільноживучих прісноводних інфузорій: темп пульсації не перевищує 1 хв. (20-30 сек.).
У Radiophryaз морської олігохети Enchytraeus sp., що мешкає на літоралі Баренцева моря, пульсація відбувається через 6-8 хв.
Нарешті, у Metaradiophrya lumbriciі Anoplophrya lumbriciз кишечника грунтової олігохети Eisenia foetidaі у Mesnilella fastigataі Radiophrya prolijeraз ґрунтових Enchytraeus sp. темп пульсації повільніший (1.5-4 хв.), ніж у інфузорій з прісноводних, і кілька більш швидкий, ніж у Astomataз морських олигохет. Таким чином, у близьких видів інфузорій Astomataвиявляється істотне фізіологічне відмінність внаслідок пристосованості до різної в осмотическом щодо середовищі. Для інфузорій з прісноводних черв'яків изотонической середовищем є 0.4% -й рінгер, тоді як для інфузорій кишечника грунтових черв'яків Ізотон відповідає 0.75-0.8% -му розчину Рінгера, а для морських форм - 3% -му розчину.
Вплив зміни кількості солей у навколишньому середовищі на темп пульсації вакуолі було продемонстровано ще Цюльцер (Zuelzer, 1910). Вона показала, що Amoeba verrucosaпри привчанні до життя в морській водівиявляє, починаючи з концентрації солі в 0.3%, поступове уповільнення пульсації вакуолі, а коли концентрація солі досягає 1.5%, вакуоль зовсім зникає, щоб виникнути знову при поступовому розведенні культури прісною водою. Точно так же Херфс (Herfs, 1922) отримав для Paramecium,привчати до життя в розчинах NaGl і в прісній воді, такі дані:
| Вміст солі у воді (в%) | 0 | 0.25 | 0.5 | 0.75 | 1 |
| Проміжок між пульсаціями (в сек.) | 6.2 | 9.3 | 18.4 | 24.8 | 163 |
| Кількість виділеної о 1 годині рідини, виражене в обсягах тіла | 4.8 | 2.82 | 1.38 | 1.08 | 0.16 |
Взагалі кількість рідини, що виводиться через вакуолі в певний строк, за інших рівних умов пропорційно темпу пульсації і у різних Protozoaможе бути дуже різним. Наприклад, скорочувальна вакуоль Uronema nigricansпри температурі 28 ° за 2 хв. виділяє обсяг води, що дорівнює об'єму тіла інфузорії, а у Euplotes patellaдля цього потрібно 14 хв., у Paramecium aurelia- 46 хв.
2) Фізіологічне значення скорочувальних вакуолей
Функції вакуолей, мабуть, різноманітні. Перш за все є підстави думати, що на частку вакуолей падає виведення з тіла нагромаджувати в ньому кінцевих продуктів обміну речовин.
Припущення про цю функцію висловлювалися ще в минулому столітті Штейном (Stein, 1878) і іншими дослідниками, але Хоуланд (Howland, 1924а, 1924b) перша побічно показала, що найпростішими виділяється сечова кислота. В її культурах парамецій і амеб була сечова кислота, концентрація якої збільшувалася приблизно пропорційно терміну утримання культури. M. Новиков (1908), Шумву (Shumway, 1917) і флатер (Flather, 1919) показали, що при згодовуванні інфузоріям щитовидної залози, при дії адреналіну і екстракту епіфіза пульсація вакуолей частішає, а самі вакуолі збільшуються, т. Е. Виходить стимуляція екскреції під впливом тих же засобів, які у хребетних викликають діурез.
Везербі (Weatherby, 1927) виявив сечовину в рідини скорочувальної вакуолі Spirostomum,видобутої за допомогою микропипетки. Однак, судячи з підрахунками цього автора, вакуоль виводить лише 1% всієї виділеної масовими культурами Spirostomumсечовини.
Загалом дані про характер азотистих продуктів виділення вельми суперечливі. Крім сечової кислоти і сечовини, деякими авторами замість цих речовин в якості продуктів виділення вказувався аміак (наприклад, у Glaucomaі Spirostomum). He зовсім узгоджується з екскреторної функцією скоротливих вакуолей лише ту обставину, що вони ніколи не беруть участь у виведенні назовні інтравітальних фарб, введених в плазму, які дуже часто виводяться у вищих тварин через нирки та інші видільні органи.
Неодноразово висловлювалася думка, що скоротливі вакуолі приймають відоме участь в дихальному обміні найпростіших, сприяючи виведенню з цитоплазми вугільної кислоти; однак фактичних доказів специфічної діяльності вакуолі в цьому напрямку немає. Тільки непрямі дані Людвіга (Ludwig, 1928) про відповідність дійсного кількості виведеної через вакуоль і можливого кількості виділеної тваринам вугільної кислоти тлумачаться цим автором на користь дихальної функції вакуолі.
Більшістю авторів в даний час висувається на перший план та важлива роль, яку скоротливі вакуолі грають в якості осморегуляторной апарату. Ще Деген (Degen, 1905) вказав, що плазма прісноводних найпростіших завдяки присутності в ній солей має вищий осмотичним тиском, ніж довкілля; а ця обставина веде до постійного вбирання цитоплазмой води. Для видалення безперервно надходить в плазму надлишку води і служить скорочувальна вакуоль. Правильність цього погляду підтверджується як характером поширення скорочувальних вакуолей серед найпростіших, так і дослідами привчання прісноводних найпростіших до води зі зростаючою концентрацією солей (Firiley, 1930; Loefer, 1939, і ін.).
нещодавно з'явилося експериментальне дослідження Jl. H. Серавін (1958) над пульсацією вакуолі Paramecium саіdatum,в якому наводяться дані, що показують, що при перекладі інфузорій в сольові розчини підвищеної концентрації у них протягом перших 15- 30 хв. спостерігається зниження швидкості пульсації вакуолі в кілька разів, потім темп пульсації поступово прискорюється і наближається до швидкості, властивої контрольним інфузоріям, але зазвичай не досягає його рівня. Наприклад, в 0.2% NaCl через 30 хв. темп пульсації був в середньому 40.5 сек., а через 48 год. досяг 13.1 сек., тоді як у контрольних пульсація протікала зі швидкістю 10.3 сек. Те ж саме спостерігалося і в розчинах 0.1% LiCi, 0.5% CaCl 2, в 1% -й сахарозі, в розчині сечовини. Уповільнення відбувалося в 7-15 разів, але через 2 доби
темп пульсації в цих розчинах був уповільнений всього лише в 1.5-3 рази. У більш низьких концентраціях солей спостерігалося повне відновлення темпу пульсації вакуолі. Такі речовини, як HGl, NaOH, GaGl 2, формалін, в незначних концентраціях викликали слабке уповільнення пульсації, яка потім швидко приходила до норми. Адреналін в концентрації 0.0005% навіть збільшив водообмін, по згодом пульсація також повернулася до норми. Таким чином, осмотично активні і неактивні речовини викликають подібні зміни діяльності скорочувальної вакуолі. При цьому спостерігається загальна тенденція регуляції водного потоку, що проходить через цитоплазму інфузорії, шляхом зміни темпу пульсації вакуолі. Крім того, відбувається пристосування інфузорій до постійно змінюваних умов осмотичного тиску зовнішнього середовища. Це ж відзначав і Кітчінг (Ritching, 1952) щодо Саrchesiumі Podophrya.У цих інфузорій в високих концентраціях етиленгліколю і сахарози відбувається повна зупинкадіяльності вакуолі, але через деякий час пульсація вакуолі і водообмін відновлюються.
Все це свідчить про складному механізмі дії скорочувальної вакуолі, що грає безсумнівну роль в водообмене інфузорій. Серавін вважає, однак, на підставі того, що осмотически неактивні речовини викликають затримку пульсації вакуолі, можливим припустити, що водообмін у інфузорій не визначається осмотическими закономірностями і тому роль скорочувальної вакуолі в осморе-регуляції не може вважатися доведеною.
Для розуміння функції вакуолі необхідно, звичайно, зрозуміти також і механізм циркуляції води з цитоплазми в резервуар вакуолі і механізм скорочення самої вакуолі. І. Гелей та Г. Гелей (Gelei, 1928; Gelei, 1939), як і ряд інших дослідників, вважають, що тільки тургорного напругою, що створюються в цитоплазмі, і, отже, осмотическими умовами визначається скорочення резервуара вакуолі. Можливо, що мається і активний механізм секреції води в вакуоль (Kitching, 1956). На думку Шнейдера (Schneider, 1950), пульсація вакуолі обумовлена діяльністю скоротних фібрил, розташованих в стінці вакуолі, а наповнення каналів, що приводять водою відбувається наступним чином. Рідина збирається в канальця ергастоплазма і потім внаслідок більшого діаметру цих канальців, ніж трубочок нефрідіальной плазми, створюється струм води з канальців ергастоплазма в трубочки нефрідіальной плазми і з них вже в просвіт каналу. Потрібні, звичайно, подальші дослідження, щоб більш точно розібратися в механізмі цих процесів. Екскреторна і дихальна діяльність скорочувальної вакуолі здаються вельми можливими, але не остаточно доведеними.
3) Скорочувальна вакуоль і апарат Гольджі
Питання про морфологічному значенні скорочувальної вакуолі вступив в нову фазу після робіт Д. H. Насонова (1924, 1925), який висловлює думку про те, що скорочувальна вакуоль найпростіших гомологична апарату Гольджі в клітинах Metazoa.Насонов (1924) і майже одночасно з ним І. Гелей (J. Gelei, 1925а, 1925b), застосовуючи техніку осмірованія до інфузорій, виявили важливі, нові риси в будові скорочувальних вакуолей (рис. 127). Насонов знайшов, що навколо скорочувальної вакуолі багатьох інфузорій (Campanella, Lionotus, Parameciuin, Nassula),а також у Chilomonasзнаходиться сильно Черняєв від
відновленого осмію оболонка, з гладкою поверхнею або оточена шаром дрібних, осміофільние зерен. Оболонка або оточує всю вакуоль цілком (у перерахованих вище видів), або ж має вид сітчастого кільця, оперізувального вакуоль по її екватору (Chilodon, Dogielella).У форм, що володіють системою каналів, що приводять (Paramecium),оболонка продовжується і на останні; при цьому канали можна простежити в ендоплазме набагато далі, ніж при будь-якому іншому методі дослідження, і дистальні частини їх оточені сітчастої муфтою сірого кольору. Цей відділ каналів Насонов вважає власне екскреторної частиною апарату, тоді як проксимальную частина каналів і саму вакуоль лише вивідний частиною вакуолярної системи. У DogielellaНасонов міг констатувати, що під час систоли вакуолі навколишній її чорне кільце не слід за останній при її скороченні, але залишається на своєму колишньому місці. Гелів (Gelei, 1925a, 1925b) пробував порівнювати їх з нефрідіальной системою Metazoa,прирівнюючи саму скоротливу вакуоль сечового міхура, інші частини апарату - іншим розділах нефридії. Насонов зупинився на схожості осмірующейся оболонки вакуолей з апаратом Гольджі у Metazoa.Дійсно, наведені ним малюнки різних типівскорочувальних вакуолей і апаратів Гольджі, взятих з різних типів клітин у Metazoa,надзвичайно переконливі по своїй схожості.
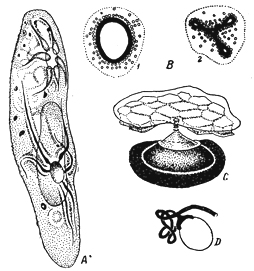
На підставі цього Насонов вважає можливим шукати в осміофільние губчастої оболонці призводять каналів у Parameciurnі сітчастого кільця у Dogitlellaне тільки морфологічного, але і функціонального паралелізму між цими утвореннями і апаратом Гольджі. Сітчастість речовини кільця залежить від наявності в ньому маленьких освітніх вакуолек - стадія пов'язаного секрету. Ці вакуолькі потім, зливаючись, утворюють центральну вакуоль, Що лежить в центрі кільця - стадія вільного секрету, що виділяється назовні. За концепцією Насонова, апарат Гольджі служить для збирання з плазми відомих речовин, які підготовляють в ньому для свого виведення геть із тіла через вакуоль. Вся фактична сторона знахідок Насонова цілком підтвердилася пізнішими дослідженнями ряду авторів: А. А. Стрєлкова (1939) у Cycloposlhiidae,Форе-Фремо (Fauré-Fremiet, 1925) у Vorlicella,Кінг (King, 1928) у Paramecium trichiumі Euplotesі ін. Правда, у деяких найпростіших не вдавалося виявити навколо вакуолі особливої осміофільние оболонки, що могло залежати від методичних моментів.
в клітинах всіх Metazoaу вигляді сітчастого утворення або що складається з окремих диктиосом. Тому можна думати, що він є також і у всіх одноклітинних організмів. Дійсно, дослідження Хіршлер (Hirschler, 1927), Овас (Hovasse, 1937), Жуае-Лавернь (Joyet-Lavererie, 1926) та інших показали наявність в плазмі багатьох найпростіших (Gregarina, Coccidia, Flagellata, Sarcodina)дрібних паличок, колечок, кульок з сильно осміофільние краєм і дуже схожих з діктіосоми клітин Metazoa.
Ознаками апарату Гольджі у Metazoaє перш за все здатність цього апарату специфічно відновлювати деякі сполуки важких металів - срібла і осмію, що виражається в почорнінні структур апарату. Ці структури не фарбуються прижиттєво нейтральним червоним і метиленовим синім. Нерідко характерними структурами апарату Гольджі є діктіосоми, які складаються з двох частин - хромофільной і хромофобная. Зазвичай секреторні гранули занурені в хромофобная частина. Як правило, апарат Гольджі локалізується саме в секреторною зоні клітини.
Дослідження з електронним мікроскопом показали єдність структур апарату Гольджі в різних клітинах Metazoa. Bo всіх досліджених клітинах виявлені подвійні мембрани (або γ-цитомембрани), розташовані паралельно один одному і утворюють систему з 3-7 і більше пар. Кожна пара обмежує більш світлий простір шириною 50-200 Å і утворює щось на зразок плоского мішечка. Іноді воно розширюється до великої вакуолі. Мембрани занурені в мелкозернистое основна речовина. У ряді випадків мембрани можуть оточувати різної величини вакуолі. Всі ці структури є в достатній мірі стабільними.
Істотним є той факт, що є повна відповідність між картинами апарату Гольджі, описаного за допомогою світлового мікроскопа Насоновим π іншими дослідниками в клітинах Metazoa,і сучасними зображеннями, отриманими з електронним мікроскопом.
Дальтон і Фелікс (Dalton a. Felix, 1957) відзначають також, що скорочувальна вакуоль Chlamidomonasобмежена мембранами, схожими з такими апарату Гольджі клітин Metazoa,і, таким чином, автори підтримують гіпотезу Насонова про гомології цих органел. Навколо скорочувальної вакуолі у інфузорій Campanellaі Ophridiumрозташовуються численні трубочки з щільними осміофільние стінками, які можуть бути до певної міри можна порівняти з ультраструктурами апарату Гольджі (Fauré-Fremiet et Rouiller, 1959). Наведені вище дані Шнейдера про будову нефрідіальной плазми навколо каналів, що приводять скорочувальної вакуолі Parameciiimроблять, проте, скрутним гомологізірованіе її структури з ультраструктурою диктиосом.
Ще раніше французькі автори (Duboscq et Grassé, 1933; Grassé et Hollande, 1941) також відкинули гомологию осміофільние оболонок скорочувальних вакуолей з апаратом Гольджі. Вони вважають, що осміофільние стінки вакуолей, як і самі скоротливі вакуолі, є абсолютно незалежними від апарату Гольджі утвореннями.
Необхідно відзначити, що у деяких інфузорій виявлені серпоподібні або кільцеподібні діктіосоми, як наприклад у Balantidium elongatum,поза зв'язком, з вакуолями (Villeneuve-Brachori, 1940). Нуар-Тімоті (Noirot-Timothee, 1957) виявила з електронним мікроскопом типові структури апарату Гольджі в ендоплазме деяких Ophryoscolecidae,які пов'язані з скорочувальної вакуолью.
Французькі автори гомологізіруют парабазального тіло з апаратом Гольджі (див. Стор. 115), так як це утворення особливо при дослідженні з електронним мікроскопом виявилося схожим з діктіосоми клітин Metazoa(Рис. 80).
Потрібні безсумнівно подальші порівняльні дослідження найпростіших, щоб вирішити питання про гомології тих чи інших структур з апаратом Гольджі клітин Metazoa.
Скорочувальна вакуоля, см. Вакуолі ... Науково-технічний енциклопедичний словник
Contractile vacuole сократительная вакуоль. Tип вакуолі у деяких груп протистов, що бере участь у виведенні води (розчинів) з клітки при скороченні і в поглинанні води клітиною при розширенні, що служить для регуляції осмотичного тиску. ... ... молекулярна біологіяі генетика. Тлумачний словник.
Структура еукаріотичної клітини. Вакуоль вказана під номером 10 Вакуоль одномембранних органоид, що міститься в деяких клітині і ... Вікіпедія
Або Heliozoa загін класу саркодікових (див.) Типу найпростіших (див.) Тварин. Морфологічні властивості. Відрізняються кулястим протоплазматіческім тілом, від якого в усіх напрямках, на зразок променів, відходять тонкі, ниткоподібні, ні ... ... енциклопедичний словникФ.А. Брокгауза і І.А. Ефрона
Або Ciliata загін класу наливу, або інфузорій (див.), Типу найпростіших (див.). Ресничном ІНФУЗОРІЇ. I (Aspirotricha). Значення букв: а порошіцу; al альвеолярний шар ектоплазми; ad.Z адоральний ряд вій; b дотикова щетина; cl вії; ... Енциклопедичний словник Ф.А. Брокгауза і І.А. Ефрона
- (Lobosea), клас найбільш просто організованих найпростіших над класу корненожек. Позбавлені внутр. скелета і зовніш. раковини. Форма тіла непостійна, розміри зазвичай від 20 до 700 мкм, рідше кілька більш. Форма і розміри псевдоподий характерні для ... ... Біологічний енциклопедичний словник
- (Flagellata s. Mastigophora, див. Табл. Біченосци, Flagellata) клас найпростіших тварин (Protozoa). Як і всі інші представники цього типу, вони мають тіло, що складається тільки з однієї клітини, що представляє протоплазму і ядро з ядерцем. ... ... Енциклопедичний словник Ф.А. Брокгауза і І.А. Ефрона
Або Protozoa. Зміст статті: Характеристика і класифікація. Історичний нарис. Морфологія; протоплазма з включеннями (трихоцисти, ядро, скоротливі вакуолі, хроматофори і ін.). Покрови і скелет. Рух П .; псевдоподии, джгутики і ... ... Енциклопедичний словник Ф.А. Брокгауза і І.А. Ефрона