Клітини грибів покриті. Зростання грибів та вплив зовнішніх факторів на ростові процеси
Процес збільшення розмірів клітин, а отже, і загальної маси характеризується поняттям «зростання». Він здійснюється у міцеліальних грибів шляхом розтягування внутрішньої оболонки верхівкової клітини. Необхідна умова зростання - наявність поживних речовин і надходження до зростаючої клітці цитоплазми з сусідніх предверхушечних клітин. Не менше значення мають активні процеси синтезу, в ході яких утворюються ядро, клітинні органели і елементи клітинної стінки. У дріжджових грибів зростання нових дочірніх клітин відбувається нерозривно шляхом брунькування або ділення. Ця здатність характерна і для деяких міцеліальних грибів.
Зростаючий ділянку гіфи містить підвищену кількість цитоплазми, білкових речовин, нуклеїнових кислот, ферментів, т. Е. Він найбільш активний в фізіологічному відношенні. За зростаючими клітинами розташовуються вакуолізірованние, цитоплазма яких через пори поперечних перегородок під впливом сил осмотичного тиску виштовхується до верхівкової клітці. У вакуолізірованние предверхушечних клітинах містяться запасні поживні речовини, в тому числі глікоген, використовувані для освіти оболонки.
Швидкість росту грибів, які ростуть на твердих поживних середовищах, найчастіше визначають шляхом виміру діаметра або радіуса і загальної площі колонії міцелію, а швидкість росту суперечка - по збільшенню обсягу їх біомаси при набуханні.
Ростові процеси сильно залежать від генетичних особливостей, умов культивування та дії зовнішніх чинників. Швидкість зростання різних грибів далеко не однакова. У зв'язку з цим їх поділяють на швидко зростаючі (Neurospora, Trichoderma, Muco-rales, Aspergillus), окремі представники яких можуть рости з високою швидкістю - до 3-6 мм / год, і медленнорастущие (Phellinus ріnі, Polystictus circinatus). Приріст швидкозростаючих дереворуйнівних грибів Fomes fomentarius і Pleurotus ostrealus становить 9-14 мм на добу, а медленнорастущих Piptoporus betulinus - лише 1,5-3,0. Е. В. Вітровий доведено, що швидкість росту гетерокаріонов вище, ніж гомокаріонов.
Міцелій (грибниця) - вегетативне тіло грибів і актиноміцетів (деякі дослідники, підкреслюючи бактеріальну природу актиноміцетів, називають їх аналог грибного міцелію тонкими нитками), що складається з тонких (1,5-10 мкм завтовшки у грибів і 0,5-1,0 мкм у актиноміцетів) розгалужених ниток, званих гіфами. Розвивається в субстраті і на його поверхні. Довжина міцелію грибів у природних умовах, зокрема, в лісовій підстилці, може досягати 35 км на 1 грам. Зростання міцелію відбувається апікально (тільки в вершині). У грибів розрізняють неклітинний (ценотичний) міцелій, позбавлений міжклітинних перегородок і представляє собою величезну клітку з великою кількістюядер (характерний для зигомицетов), а також клітинний (септірованний), з наявністю міжклітинних перегородок (позначаються терміном септи) і одного або багатьох ядер в окремій клітці.
Міцелій актиноміцетів не має ядер і може як розділятися на окремі клітини, так і залишатися єдиним. Септи можуть бути з простими і складними порами. Прості характерні для аскомицетов. Складні пори часто супроводжуються пряжками - гачкоподібними виростами біля септ, поєднаних з однією клітиною і придатними до сусідньої. В такому випадку клітина має два ядра і міцелій гриба називають дікаріотіческім. Він характерний для аскомицетов і базидиомицетов. Пряжки грають важливу роль при діленні клітин. Плодові тіла базидіоміцетів утворені помилковою тканиною плектенхімой, що складається з густо переплетених гіфів міцелію і відрізняється від справжньої тканини тим, що справжня формується при діленні клітин в трьох напрямках, а помилкова - в одному. У базидиомицетов міцелій часто багаторічний, у інших грибів - однорічний.
Форми міцелію:
Плівки - щільні плоскі сплетіння гіф, різних розмірів, товщини, світлих відтінків служать для прикріплення плоті до субстрату. Руйнують ферменти до целюлози і поглинають її.
Шнури - зрощені Стек гілок гіфи гриба. Для прикріплення субстрату і його поширення. Бувають коротко розгалужені або довгі сильно розгалужені.
Різоморфи - потужні шнури до 5 метрів, складаються з темних щільних зовнішніх ниток гіф і внутрішніх світлих пухких шнурів.
Ризоктонії - тонкі повітряні шнури. Служать для прикріплення і поширення.
Склероції - дуже щільні сплетіння гіф. Містять багато ниткоподібних розгалужень. Часто склероции утворюються з плодоношення гриба. Служать для полегшення перенесення несприятливих умов.
Строми - плоскі щільні зрощення з тканиною рослини господаря. Служать для збереження спор гриба.
Плодове тіло - служить для утворення споро носіння гриба.
Зростання міцелію припиняється при повному використанні живильного середовища або при переході гриба в наступні стадії розвитку, пов'язані з розмноженням. Таким чином, вегетативний ріст - підготовчий етап, який створює сприятливі умови для розмноження. Мікроскопічні дослідження міцелію дозволили виділити у грибів ряд вікових стадій, що характеризуються певними ознаками.
I стадія - проростання конідій (суперечка). Включає два етапи: перший - набухання конідій (триває 4-10 год), другий - утворення ростових трубок і первинного міцелію з декількох клітин. Цитоплазма не диференційовані, в клітині містяться дрібні вакуолярного плями.
II стадія - розростання міцелію і його розгалуження (лагфаза). Ядро набуває характерного для грибів вид. Утворюються дрібні гранули ліпідів.
III стадія - рівномірний інтенсивний ріст, що супроводжується активними обмінними процесами і накопиченням нуклеїнових кислот.
IV стадія - сповільнене зростання, старіння міцелію, спорообразование, накопичення вторинних метаболічних продуктів.
V стадія - прийнята для умовного позначення повного автолиза міцелію.
Що стосується окремих грибам, зокрема до продуцентів пеніциліну, детально описані всі вікові зміни. Так, у Penicillium chryzogenum описано 7 вікових стадій.
Тривалість росту грибів в тій чи іншій стадії залежить від екологічних чинників, Генетичних особливостей, умов культивування та ін.
клітка грибаскладається з клітинної оболонки (зовні вона часто буває слизовим шаром-капсулою), Ломас, цитоплазми з цитоплазматичної мембраною, ендоплазматичної мережею, мітохондріями, рибосомами, діктіосоми і ядрами. Іноді в клітині грибів є вакуолі і різні включення. Клітинна оболонка, що здійснює у грибів численні функції, в тому числі активного всмоктування поживних речовин із субстрату, в якості основних компонентів містить хітин, полісахариди, в тому числі глюкани, білки і жири. У клітинній оболонці грибів є також пігменти (меланіни, хінони), сюди ж входять різні іони і солі. Електронно-мікроскопічне вивчення оболонок клітин грибів показує, що вони складаються з декількох шарів фибриллярного будови. Ці фібрили, що представляють собою білкові мікротрубочки утворюють скелет, який служить основою для інших компонентів оболонки. Клітинна оболонка надає форму клітинам гіф та органам розмноження. Відмінними ознаками клітинної оболонки деяких представників нижчих грибів є відсутність в ній хітину і наявності тільки целюлози. В цитоплазмі, у цитоплазматичної мембрани, у грибів розташовані ломасоми - губковідние електронно-прозорі структури.
Цитоплазми грибної клітиниявляє собою рідку колоїдну середовище, в якій містяться структурні білки, клітинні організмиі не пов'язані з ними ферменти, амінокислоти, вуглеводи, ліпіди та інші речовини. Вакуолі - структури округлої, рідше неправильної форми, які виконують функцію депо для відкладення запасних речовин або ж токсичних продуктів метаболізму. В якості резервних речовин, тут запасаються в основному поліфосфати (метахроматін, волютин), глікоген, ліпіди.
мембранна системапредставлена ендоплазматичної мережею у вигляді розгалужених в цитоплазмі і пов'язаних між собою мембранних канальців, цистерн і порожнин, що виконують функцію внутрішньоклітинної та міжклітинної транспортної мережі для метаболітів.
ядроокруглої або подовженої форми, оточене подвійною мембраною, має ядерце і хромосоми з ДНК. Кількість ядер в грибний клітціі їх розміри різні. Відомі як одноядерні клітини, так і клітини, кількість ядер, в яких досягає декількох десятків; розміри ядер також коливаються від 2-3 мкм в діаметрі до декількох десятків мікрометрів. Для грибів, яким властива дікаріотіческая фаза в розвитку. Характерно наявність двох ядер, спарених у вигляді дикаріон. Також грибам характерні всі інші органи тваринної клітини.
Вегетативне тіло грибівскладається з гіф, що мають вигляд циліндричних трубок до 10 мкм в діаметрі, вони мають верхівковим (апікальним) зростанням і рясним розгалуженням. Всередині, гіфи виконані протоплазми; у вищих грибів є поперечні перегородки і утворюються вони зазвичай на певній відстані від кінця гіфи. Значного різноманітності досягає будову клітинних перегородок, або верес, які є похідними клітинної оболонки і утворюються шляхом інвагінації (випинання) цитоплазматичної мембрани всередину клітини. Це властивий всім грибам спосіб виникнення сент. Через них здійснюється зв'язок з цитоплазмою сусідніх клітин, відбувається переміщення поживних речовин, міграція деяких клітинних органів. Для більшості базидиомицетов характерний доліпоровий тип, що мають складну будову. Гіфи вищих грибів, сплітаючись між собою, утворюють міцелій, у окремих видів він створює подобу тканини.
гриби розмножуютьсявегетативним безстатевим або статевим способами. вегетативне розмноженняздійснюється фрагментами міцелію, які, відділяючись, дають початок новому міцелію. У дріжджових грибів і представників порядків Agaricales і Plectascales відомо вегетативне розмноження шляхом брунькування міцелію або його клітин, в результаті чого утворюються окремі клітини-іодіі, що дають початок грибного організму. Для цілого ряду грибів характерно вегетативне розмноження шляхом розпаду на окремі клітини-артроспори.
при безстатевому розмноженніспори набагато більш високо спеціалізовані за будовою і способи розмноження. Серед суперечка безстатевого розмноження грибів за способом освіти виділяють суперечки ендогенні і екзогенні.
статеве розмноженняу грибів буває різних типів. Сутність його полягає в тому, що відбувається злиття двох статевих клітин (гамет) - чоловічої і жіночої - або двох вегетативних талломов, що функціонують як статеві клітини, в результаті виникає новоутворення (зигота). Зливаються гамети містять тільки половинний набір хромосом. У зиготі число хромосом відповідно подвоюється. Гамети є структурами, які знаходяться, маючи половинний хромосом, в гаплоидной фазі, а зигота переходить вже в диплоїдну фазу.
У вищих грибів статевий процес протікає як злиття органів і клітин, що не диференційованих на гамети. Новоутворена в результаті злиття зигота (теж не диференційована і зазвичай представляє собою лише відповідне ядерне стан) без періоду спокою переходить до подальшого розвитку; в ній формуються дікаріони ядер протилежних статей, які потім попарно зливаються і зазнають редукційний розподіл. Гаплоїдні ядра, які утворилися в процесі редукційного поділу, переходять в аскоспори, що утворюються в сумках або в базидіоспори, що утворюються на спеціальних клітинах - базидіях - базидіоміцети екзогенно.
Гриби поширені повсюдно: їх суперечки, обривки міцелію, інші освіти, зустрічаються на грунті і в повітрі, на суші в воді. Вони розвиваються на всіляких природних субстратах рослинного і тваринного походження, а також на штучних матеріалах створених людиною. У XX ст. перед людством постала проблема збільшення природних і штучних джерел білка, дефіцит якого стає все відчутнішою. У зв'язку з цим виникла необхідність введення в культуру нових белоксодержащіх організмів, серед яких одним з найбільш цінних є їстівні гриби. Культивування їстівних грибів дозволяє запобігти харчові отруєння, викликані споживанням дикорослих грибів. Вирощувати їстівні гриби можна цілий рікнезалежно від кліматичних і ґрунтових умов, На поживних субстратах, малопродуктивних для інших цілей, наприклад на різних не харчових відходах; при цьому субстрат зазвичай використовується двічі: після збору врожаю грибів він стає цінним джерел перегною для садівництва і овочівництва. Підвищення попиту на гриби на світовому ринку сприяло подальшому вдосконаленню методів їх вирощування на основі глибокого вивчення біології культури.
У наших дослідженнях ми досліджували гливу звичайну. Вона входить до збірної групу макроміцети (макро - великі, міцети - гриби). За будови вегетативного тіла макроміцети належать до вищих грибів. Їх міцелій багаторічний. Поселившись на певному субстраті, він виростає нерідко на багато метрів в довжину. У міру зростання гіфи гілкуються, переплітаються. У місцях їхнього зіткнення виникають перемички (анастомози); ці перемички об'єднують гіфи в єдиний організм, здійснюють зв'язок між ними, передачу поживних речовин.
Дереворазрущающій міцелій Гливи розвивається в повітряний міцелій, схожий на пишні шматочки вати.
Міцелій здійснює всі життєво важливі функції грибного організму - його харчування, ріст і розвиток, розмноження. За способом харчування макроміцети, як і інші гриби, гетеротрофи, такт як позбавлені здатності до фотосинтезу. Тому вони живуть тільки там, де вже є готове органічна речовина, і добувають його з найрізноманітніших джерел.
Накопичивши достатній запас поживних речовин, грибниця стає здатною до розмноження. У макроміцетов цей процес пов'язаний з утворенням грибного тіла - тієї частини грибного організму, яку ми зазвичай називаємо грибами, забуваючи або зовсім не знаючи про те, що це лише органи розмноження, що виникають на певному етапі і призначені для розвитку спор і їх захисту. Плодові ж тіла різноманітні, розташовуються, як правило, на поверхні субстрату - отже, їх зручно розглядати і вивчати. Складні плодові тіла макроміцетов оточені грибний помилкової тканиною, або плектенхіми, яка складається з більш-менш щільного сплетіння гіф.
Гриби і рослини мають такі подібності - надходження поживних речовин в клітину, засноване, головним чином, на явищах осмосу (дифузії речовин через напівпроникні перегородки поверхні клітин), не є чисто фізичним, але є фізіологічним явищем. При надходженні поживних речовин в клітину гриба, клітина відіграє роль активну, а не пасивну, так як проникність протоплазми, від якої вона залежить, є величиною змінною. Крім того, існує виборча проникність для певних речовин і при тому різна в різному стані клітин організму. Звідси з'являється можливість застосування подібним чином деяких стимуляторів, наприклад гетероауксину і епін для грибів.
Дікаріотная грибниця здатна на необмежене зростання - вегетативне тіло грибів складається з гіф мають вигляд циліндричних трубок до 10 мкм в діаметрі, вони характеризуються верхівковим (апікальним) і необмеженим ростом і рясним розгалуженням.
Відсутність здебільшого рухливості в вегетативному стані у вищих рослинтканинне будова виникає при діленні клітин у всіх напрямках. А у грибів міцелій ділиться тільки з утворенням поперечних напрямків, тобто тільки в одному напрямку. Тому прийнято вважати, що у грибів немає справжніх тканин, а є лише помилкові тканини. Залежно від морфологічних особливостей у грибів розрізняють два типи тканин: параплектенхіму і прозоплектенхіму.
Крім морфологічного поняття існує і фізіологічне поняття тканини у грибів. З точки зору функціонального призначення розрізняють покривні, механічні та провідні тканини. з покривної тканинискладається поверхню склероциев і плодових тіл вищих грибів. Клітини такої тканини мають потовщені оболонки, на поверхні відкладається пігмент, який поглинає промені сонячного спектра, тим самим виконує захисну роль. Механічна тканина представлена гіфами сильно потовщеними стінками і судженим просвітом, які надають міцність плодового тіла або будь-якої його частини. Типовою провідної тканини у грибів немає, його функції виконують особливі спеціалізовані гіфи позбавлені поперечних перегородок. Ці гіфи, пронизуючи плодове тіло в різних напрямках, забезпечують його водою. Для просування органічних речовинє гіфи, які є відгалуженнями звичайних гіф. Вони відрізняються густим пофарбованим вмістом.
Все перераховане - тобто функціональне схожість тканин вищих рослин і спеціалізованих гіф грибів говорить про ще один схожості.
Вегетативне розмноження - вегетативне розмноження здійснюється фрагментами міцелію, які, відділяючись, дають початок новому міцелію. У агарикових відомо вегетативне розмноження шляхом брунькування міцелію або його клітин, в результаті чого утворюються окремі клітини - Оїдії, що дають початок новому грибного організму.
Їстівні гриби характеризуються помітним кількістю мінеральних речовин, вміст яких у окремих видів може досягати 11,5% (в середньому 7,7%). Плодові тіла грибів багаті калієм, фосфором, в незначних кількостях міститься в них натрій і кальцій, вміст заліза у макроміцетов приблизно аналогічно тих кількостей його, яке виявляють у більшості рослинних продуктів.
Тобто в наявності схожість в хімічному складі плодових тіл вищих грибів і продуктів вищих рослин. А тепер ще раз більш схематично про розмноження грибів.
розмноження грибів
У макроміцетов розрізняють статевий, безстатеве і вегетативне розмноження. Вегетативне розмноження, в основі якого лежить здатність організму до регенерації, може здійснюватися за допомогою частин міцелію. Здатність грибів до вегетативного розмноження широко використовується при штучному розмноженні культивованих грибів, а також при пересіву чистих культур в лабораторіях, які виробляють посівної міцелій.
Більш спеціалізованим вважається спосіб, при якому міцелій розпадається на окремі клітини, які згодом проростають, утворюючи міцелій. До органів вегетативного розмноження відносяться Оїдії, хламідоспори, які, хоча і порівняно рідко, утворюють деякі види вищих грибів. Хламідоспори - це товстостінні ділянки гіф, що відокремилися від міцелію і покриті темною, щільною оболонкою. Вони здатні зберігати життєздатність від року до десяти років. При настанні сприятливих умов вони проростають в новий міцелій. Оїдії - короткі циліндричні тонкостінні ділянки гіф, утворені в результаті повного розпаду міцелію при вологих умовах. При проростанні дають початок новому міцелію.
Для багатьох макроміцетов характерно і безстатеве розмноження. Воно здійснюється за допомогою спеціалізованих клітин або багатоклітинних структур (суперечка), які проростають в міцелій.
Не зупиняючись на різноманітності форм статевого розмноження грибів, розглянемо статевий процес у базидіальних грибів, який називається соматогамия. Він складається в злитті двох клітин вегетативного міцелію шляхом утворення анастомозів між нитками міцелію або іншим шляхом і освіти дикуни-отіческого міцелію, на якому утворюються базидії з базидиоспорами. Базидіоспори гаплоидни, вони дають початок гаплоидному міцелію, і все починається спочатку.
Міцелій вищих грибів, будучи аеробних організмом, зростає і розвивається в природних умовах вегетативним шляхом, уникаючи надмірно вологих і заболочених місць. При певних умовах зовнішнього середовища він досягає біологічної зрілості і утворює репродуктивні органи - плодові тіла. У них розвиваються суперечки, які служать для розмноження і поширення виду в природі статевим шляхом. При відсутності умов для утворення плодових тіл міцелій вищих грибів продовжує рости і розвиватися тільки вегетативним шляхом за допомогою подовження і розгалуження гіф, а у деяких видів утворюються оідіоспори і хламідоспори.
Цикл життя гриба
Гаплоїдні базидіоспори - Haploid basidiospores проростають у відповідній навколишньому середовищуі перетворюються в недовгий гаплоїдний міцелій - haploid mycelia.
Гіфи - hyphae від двох протилежних типів гаплоидного міцелію - haploid mycelia спаровуються, зливаючи свої плазми - plasmogamy.
Створюючи дікаріотний міцелій - dikaryotic mycelium, який зростає швидше, ніж батьківський гаплоїдний міцелій - haploid mycelium, і, в кінцевому рахунку, заповнює собою весь простір. Міцелій - Mycellium гриба, ілюстрований тут (Cortinarius) формує мікоризу - mycorrhizae з деревами. Фактори навколишнього середовища типу дощу, температурних змін, і, для mycorrhizal різновиди, сезонні змінив господаря - рослині грають величезну роль в житті гриба.
Дікаріотний міцелій - dikaryotic mycellium формує компактні маси, які розвиваються в гриби. Цитоплазма - Cytoplasm, струмлива в міцелії - mycellium і також, притікає від мікоризи - mycorrhizae, роздуває гіфи - hyphae грибів, змушуючи їх "вискочити" раптово. Не випадково, з арабського, гриби називаються "Футарі" - вилазити назовні, прорізати, розколювати, раптово виявлятися назовні, схоже з російським "ранок" (гриби збирають рано вранці).
Дікаріотний базидіоміцети - Dikaryons basidiomycetes довговічний, взагалі виробляючи новий урожай - плодове тіло - basiocarps (гриби, в цьому випадку) щороку.
Karyogamy - злиття клітин відбувається в терміналі дікаріотной - dikaryotic осередки, які покривають поверхні зябер.
Кожна осередок роздувається, щоб формувати диплоїдний базидий - diploid basidium, який швидко піддається мейозу - meiosis і утворює чотири гаплоїдних - haploid ядра.
Базидий - Basidium тоді вирощує чотири придатка, і одне гаплоидное - haploid ядро входить в кожен придаток і розвивається в базидіоспори - basidiospore.
Коли дозріють, базидіоспори - basidiospores просуваються злегка (електростатичними силами) в місця між зябрами - пластинками капелюшки. Після того, як суперечки висипаються з капелюшка. Вони розсіюються вітром. Причому в капелюшку гриба дозріває від сотні мільйонів до мільярда суперечка. Це при тому, що для утворення дікаріотного міцелію потрібні всього дві суперечки.
Ці біологічні особливості вищих грибів характерні і при культивуванні в штучних умовах.
Фізіологія вищих грибів служила об'єктом вивчення ще в минулому столітті, але більш глибокі дослідження почалися в останні два-три десятиліття, коли було відкрито, що деякі цвілі мають антибіотичні властивості, а інші можуть служити джерелом ряду органічних сполук (Фостер, 1950).
Ріст і розвиток кожного окремого організму знаходяться в прямій залежності від типу поживного субстрату, в якому відбуваються складні фізіологічні та біохімічні процеси, їх інтенсивність визначається спадковими і потенційними якостями самого організму і факторами зовнішнього середовища. До факторів, що визначаються самим організмом, відносяться вид і штам гриба, походження, вік культури, кількість посівного матеріалу, здатність до вегетативного розмноження і, утворення біологічно активних речовин, інтенсивність дихання і ін.
При штучному культивуванні їстівних грибів крім факторів, придбаних організмом в процесі еволюції, необхідно враховувати і регулювати фактори зовнішнього середовища, що впливають на фізіологічні та біохімічні процеси, що відбуваються в субстратах, на формування плодових тіл, і врожайність культури.
До найважливіших факторів, що визначають активність гетеротрофних організмів, слід віднести наявність в середовищі елементів живлення, створення оптимальних умов температури, вологості, світла, реакції середовища.
Зростання грибів та вплив зовнішніх факторів на ростові процеси
Зростання грибів. Процес збільшення розмірів клітин, а отже, і загальної маси характеризується поняттям «зростання». Він здійснюється у міцеліальних грибів шляхом розтягування внутрішньої оболонки верхівкової клітини. Необхідна умова зростання - наявність поживних речовин і надходження до зростаючої клітці цитоплазми з сусідніх предверхушечних клітин. Не менше значення мають активні процеси синтезу, в ході яких утворюються ядро, клітинні органели і елементи клітинної стінки. У дріжджових грибів зростання нових дочірніх клітин відбувається нерозривно шляхом брунькування або ділення. Ця здатність характерна і для деяких міцеліальних грибів.
Зростаючий ділянку гіфи містить підвищену кількість цитоплазми, білкових речовин, нуклеїнових кислот, ферментів, т. Е. Він найбільш активний в фізіологічному відношенні. За зростаючими клітинами розташовуються вакуолізірованние, цитоплазма яких через пори поперечних перегородок під впливом сил осмотичного тиску виштовхується до верхівкової клітці. У вакуолізірованние предверхушечних клітинах містяться запасні поживні речовини, в тому числі глікоген, використовувані для освіти оболонки.
Швидкість росту грибів, які ростуть на твердих поживних середовищах, найчастіше визначають шляхом виміру діаметра або радіуса і загальної площіколонії міцелію, а швидкість росту суперечка - по збільшенню обсягу їх біомаси при набуханні.
Ростові процеси сильно залежать від генетичних особливостей, умов культивування та дії зовнішніх чинників. Швидкість зростання різних грибів далеко не однакова. У зв'язку з цим їх поділяють на швидко зростаючі (Neurospora, Trichoderma, Muco-rales, Aspergillus), окремі представники яких можуть рости з високою швидкістю - до 3-6 мм / год, і медленнорастущие (Phellinus ріnі, Polystictus circinatus). Приріст швидкозростаючих дереворуйнівних грибів Fomes fomentarius і Pleurotus ostrealus становить 9-14 мм на добу, а медленнорастущих Piptoporus betulinus - лише 1,5-3,0. Е. В. Вітровий доведено, що швидкість росту гетерокаріонов вище, ніж гомокаріонов.
Зростання міцелію припиняється при повному використанні живильного середовища або при переході гриба в наступні стадії розвитку, пов'язані з розмноженням. Таким чином, вегетативний ріст - підготовчий етап, який створює сприятливі умови для розмноження. Мікроскопічні дослідження міцелію дозволили виділити у грибів ряд вікових стадій, що характеризуються певними ознаками.
I стадія - проростання конідій (суперечка). Включає два етапи: перший - набухання конідій (триває 4-10 год), другий - утворення ростових трубок і первинного міцелію з декількох клітин. Цитоплазма не диференційовані, в клітині містяться дрібні вакуолярного плями.
II стадія - розростання міцелію і його розгалуження (лагфаза). Ядро набуває характерного для грибів вид. Утворюються дрібні гранули ліпідів.
III стадія - рівномірний інтенсивний ріст, що супроводжується активними обмінними процесами і накопиченням нуклеїнових кислот.
IV стадія - сповільнене зростання, старіння міцелію, спорообразование, накопичення вторинних метаболічних продуктів.
V стадія - прийнята для умовного позначення повного автолиза міцелію.
Що стосується окремих грибам, зокрема до продуцентів пеніциліну, детально описані всі вікові зміни. Так, у Penicillium chryzogenum описано 7 вікових стадій.
Тривалість росту грибів в тій чи іншій стадії залежить від екологічних факторів, генетичних особливостей, умов культивування та ін.
Вплив зовнішніх факторів на ростові процеси. Умови навколишнього середовища - температура, рН середовища, світло, аерація, вологість - протягом усього життєвого циклу грибів постійно роблять свій вплив. Перш за все вони позначаються на характері та особливостях ростових процесів, т. Е. Швидкість росту - один з провідних показників відповідної реакції грибів на вплив зовнішніх чинників.
Температурні чинники. Достатня кількість тепла - одне з основних умов життя гриба. Причому активна життєдіяльність кожного виду можлива в межах певних температурних меж. Найбільш гармонійний перебіг обмінних процесів спостерігається при температурному оптимумі. Вплив дуже низьких або дуже високих температур призводить до глибоких порушень в координації процесів життєдіяльності, до незворотних ушкоджень цитоплазми. Якщо температура знижується і переходить в критичну, припиняється рух цитоплазми, втрачається напівпроникливості мембран і клітина гине. Висока температура також призводить до загибелі клітини внаслідок "порушення мембран, що настає в результаті інактивації і денатурації білків і розлади обмінних процесів.
Нижня межа, при якому припиняється ріст клітин абсолютної більшості грибів, - 0-3 ° С, а верхній - не перевищує 40 ° С. Єдиного температурного оптимуму для грибів не існує, так як потреба в температурі для росту міцелію, утворення плодових тіл і проростання спір не однакова. Наприклад, оптимум температури для проростання спор у забірного трутовика Qloephillurn seriarium становить 30-34 ° С, а для росту грибниці - 36 ° С.
Зниження інтенсивності росту або повна відсутність його у грибів при певній температурі в деяких випадках викликано нездатністю синтезувати необхідні амінокислоти і вітаміни. У дослідах Д. Фріза культури Coprinus fimetarius відрізнялися гарним ростом навіть при температурі 44 ° С, якщо в середовищі містився гідролізат казеїну, активним компонентом якого є метіонін. Очевидно, блокування синтезу цієї амінокислоти при підвищеній температурі спричиняє припинення росту міцелію. Залежно від потреби в температурі гриби поділяються на три групи: холодолюбиві, тепловинослівие і теплолюбні.
Відсутність зростання у грибів при температурі нижче нуля не означає, що вони гинуть. Деякі плодові тіла сімейства Polyporaceae здатні витримувати низькі температури до -100 ° С, а міцелій Lentinus lepideus, Fomitopsis carnea виносить сухий жар до + 100 ° С, хоча зволожене повітря викликає відмирання грибниці ще при 55 ° С. Разом з тим плодові тіла, які мають м'ясисту або м'яку шкірясту консистенцію і відносяться до однорічним (Роlyporus, Polystictus і ін.), взимку гинуть від морозів. У свою чергу, дерев'янисті і пробкові плодові тіла грибів з родів Fomes, Daedalea і інші відрізняються високою стійкістю до низьких температур і з настанням весни знову починають вегетувати.
Показовими екологічні модифікації різного географічного походження. Так, середньодобовий зростання Fusarium sporotrichioides у північних культур (якутських) зазвичай нижче, ніж у південних ізолятів (азербайджанських і краснодарських).
Можливість розвитку кожного гриба, перш за все, визначається рН середовища, або реакцією субстрату в природних умовах. Живильне середовище, що містить всі необхідні речовини, але приготовлена без урахування необхідної кислотності (або лужності), може зовсім виключити можливість розвитку організму або не зробить на нього сильного інгібуючої дії. Це пояснюється утворенням аміаку і органічних кислот, що підвищують рН, внаслідок неоднакового поглинання компонентів середовища, засвоєння катіонів та аніонів. При фізіологічних дослідженнях важливо зберігати початкове значення рН середовища, використовуючи крейда, луги або фосфатний буфер.
Оптимальне значення рН для більшості грибів нижче 7 (в межах 5,0-6,0), що відповідає слабокислою реакції середовища. Разом з тим є гриби, пристосовуються до субстрату з більш кислою реакцією. До них відносяться дереворазрушающие, підстилкові і мікоризні гриби, що утворюють органічні кислоти. М. І. Бойко, який визначив зростання Hetero-basidion annosum у 9 штамів гриба, показав, що їх зростання може здійснюватися в діапазоні рН від 2,5 до 9,5, причому накопичення біомаси при рН від 5,5 до 9,5 зменшується . У деяких представників мікоризних грибів роду Boletus оптимум рН знаходиться в межах 3.
Показано, що рН середовища впливає на активність протеаз. Так, у гриба Emericellopsis glabra при зміні значення рН реакційної суміші від 6,0 до 12 активність протеаз збільшується в 5,6 рази.
Зростання грибів на поживних середовищах в більшості випадків супроводжується зміною рН в сторону підкислення. Оригінальні досліди були проведені І. Г. Коневской за визначенням зміни рН середовища, в якості якої була взята папір (сульфатна, лляна, сульфітна, бавовняна і газетна), інокулював целлюлозоразрушающіх грибом Chaetomium globosum. Після 60 діб досвіду рН сульфатної, бавовняної, лляної і сульфітної папери змінилася в бік підкислення, а газетної - в сторону подщелачивания.
Світлові фактори і випромінювання. Сонячна радіація робить істотний вплив на процеси життєдіяльності грибів, хоча дія різних ділянок спектра сонячного випромінювання неоднаково. Так, довгохвильове випромінювання викликає активацію теплових рецепторів, мутагенний ефект роблять ультрафіолетові промені, а видиме світло впливає на фотозахисні і фотохімічні процеси. Більшість грибів росте з приблизно однаковою інтенсивністю на світлі і в темряві. Однак під впливом яскравого світла спостерігається пригнічення росту міцелію і проростання спор у представників Trichoderma, Penicillum і інших грибів, особливо з безбарвними оболонками. Вельми значно світло діє на формування органів плодоношення. Плодові тіла Heterobasidion annosum, Polystictus abietinus, Armillariella mellea і багатьох інших вищих грибів при вирощуванні на штучних поживних середовищах утворюються тільки в умовах розсіяного освітлення. Показово, що різне дію світла на спорообразование можна виявити навіть всередині одного роду. Якщо Botrytis cinerea спороносит на світлі і в темряві, то В. gladiolorum - тільки на світлі. При односторонньому освітленні спостерігається фототропіческая реакція спороносних органів в сторону джерела світла. Добре помітні позитивні фототропіческіе вигини дають представники Mucorales, зокрема спорангієносцями Pilobolus cry-stallinus, види пологів Phycomyces, Spinellus, Hirschio-porus і ін. Для деяких піреноміцетов характерний вигин хоботка Перітеціі, у сумчастих аналогічним чином згинаються сумки, а у шапинкових базидиомицетов - ніжки .
Повна відсутність світла у одних грибів (Pilobolus, Lentinus, Coprinus і ін.) Викликає стерильність грибниці, в той час як у інших (Aspergillus, Me-rulius, Schizophyllum і ін.) Спороносні органи можуть розвиватися і в темряві. Нормальний розвиток плодових тіл у Lentinus edodes спостерігається при інтенсивності освітленості від 5 лк і вище. При відсутності світла утворюються светлоокрашенние гіпертрофовані плодові тіла. Під дією світла в міцелії і суперечках деяких грибів утворюються пігменти.
Слово «пігмент» походить від латинського pigmentum - фарба. Завдяки «кольоровості» пігментів вони здатні поглинати певні промені спектру. Пігментація оболонок суперечка значно підвищує їх стійкість до дії прямих сонячних променів при переміщенні повітряними течіями. Пігменти грибів поділяються на кілька груп. 1. Пігменти, що утворюються в результаті перетворення орілпірувата (терфенілхінони і їх похідні - атроментін, поліпровал, телефоровая, пульвіновая, варіегатовая кислоти і ін.). 2. Пігменти, що відбуваються по ацетатно-мевалонатного шляху (скутігерал, траметін, дермоцібін, дерморубін, дермолютеін, емодін і ін.). 3. Пігменти, що утворюються по мевалонатного шляху (лактаровіолін, лактарофульвін, Різні каротиноїди, в першу чергу (каротин, а також лакопін, кантаксантин і ін.). 4. Азотомісткі пігменти (ціннабарін, трамесангвін), а також Ряд пігментів різної структури і походження (азахіони, індиго і ін.).
Слід підкреслити, що світ не є необхідною умовою для синтезу у грибів всіх пігментів. Так, біосинтез синього пігменту індиго у гриба Schizophyljum залежить від освіти яблучної кислоти, що накопичується в міцелії в міру його розвитку. Цей пігмент утворюється лише в культурі, яка втратила здатність до синтезу яблучної кислоти. У той же час при додаванні до середовища іонів міді зміст яблучної кислоти знижується і збільшується кількість індиго.
Для деяких дріжджів, зокрема Candida pulcherria, характерний яскраво-червоний пігмент пульхеррімін.
Численну і широко поширену у нижчих грибів Mucoraceae, Choanephoraceae, Могtierellaceae і ін.) Групу пігментів становлять каротиноїди. Інтенсивним синтезом каротину мають BlakesJea trispora і Choanephora cucurditarum, які можуть бути використані для отримання 3-каротину. У дослідах Е. П. Феофілова, показано, що на синтез каротиноїдів сильно впливає дифениламин. У гриба Blakeslea trispora міцелій, відмитий від дифениламина, синтезує каротин значно швидше, ніж у контрольній культурі, що пов'язують з накопиченням безбарвного попередника, який далі використовується на синтез каротину. Якщо ж міцелій НЕ відмитий від дифениламина, то в ньому не відбувається утворення жирових включень, накопичених помаранчевими гранулами каротину. Поряд з цим змінюється і морфологія клітин міцелію. У інших грибів дифениламин викликає накопичення фітоіна і фітофлуіна, т. Е. Виявляється специфічний вплив зазначеного з'єднання на біосинтез пігментів.
Каротиноїди - похідні ізопрену, що представляють собою ненасичені сполуки терпенового ряду, належать до нерозчинним в жирах пігмеітам. Знайдено ці речовини у грибів всіх систематичних груп, хоча у багатьох вони містяться в невеликій кількості. Наприклад, у базидіоміцетів Clitocybe venutissima - і каротини складають 0,17% сухої маси плодових тел. Підвищений вміст каротину характерно для аскомицетов (порядки Taphrinales, Protomycetales і ін.) І базидиомицетов, особливо іржавинних грибів, що мають жовту і помаранчеве забарвлення. Каротиноїдні пігменти знайдені у багатьох недосконалих грибів як Темна (Cladosporium, Curvularia і ін.), Так і светлоокрашенних (Cephalosporium, Trichotecium і ін.). Роль їх в організмі грибів досить різноманітна і перш за все пов'язана з реакціями фототропізму і фототаксису.
Гриби містять багато пігментів - похідних хинона (атрометін, лейкомелон, аураутіацін і ін.), Антрахинона, бензохинона і нафтохинона. Велике число різних типів хинонов міститься у недосконалих грибів порядку гіфоміцети (Penicillium, Аlternaria, Aspergillus і ін.). Хінони мають антибіотичну і токсичною дією, пофарбовані у фіолетовий або майже чорний колір. Пігменти грибів мають найрізноманітнішу забарвлення - жовту, коричневу, червону, чорну, зелену, фіолетову і ін. Вони викликають фарбування не тільки міцелію і плодових тіл, але і живильного середовища, на якій ростуть гриби.
Багато гриби містять темні пігменти - меланіни, що представляють собою високополімерні сполуки, що утворюються при ферментативному окисленні фенолів. Меланиновой ферменти надають ряду видів грибів високу стійкість до екстремальних Умов існування.
З променів сонячного спектра найбільш сильний вплив надають ультрафіолетові, які можуть викликати мутації, а при високих дозах опромінення - повністю пригнічувати життєдіяльність грибів.
Є відомості про великий інтенсивності дії на деякі гриби синьо-фіолетових променів.
У Aspergillus clavatis при впливі синіми променями формуються видовжені конідієносци, під дією же червоних променів розмір конидиеносцев різко зменшується. Ступінь освітленості сильно впливає і на швидкість звільнення суперечка з плодових тіл. У деяких аскомицетов (Podospora curvula, Nectria cinnabarina) - спорообразование настає лише в денні години, у інших (Hypoxylon fuscum) - тільки в нічний час. У ряді дослідів показано гнітючий вплив інфрачервоних променів на зростання будинкових (Serpula lacrymans, Poria vaporaria, Coniophora cerebella) і дереворуйнівних (Heterobasidion annosum) грибів.
Сильний вплив на гриби надає іонізуюче опромінення, хоча Темна гриби виділяються своєю резистентністю до опромінення. Отримано непрямі докази того, що іонізуюча радіація викликає ушкодження ДНК. Особливою чутливістю до радіовипромінюванню мають мутанти Aspergillus nidulans, Coprinus lagopus і ін. Дози, які надають летальну дію на гриби, головним чином цвілеві, використовуються для захисту матеріалів від мікодеструкторов, порятунку мистецьких цінностей і археологічних документів.
Світло впливає на статеву і безстатеву репродукцію у грибів. Одним із з'єднань, які індукують репродуктивні процеси, є гормоноподобное речовина, що отримало назву Р310, виділене з міцелію Ascochytapisi, підданого дії ближнього ультрафіолету та інших недосконалих грибів.
Аерація. Серед грибів немає облігатних анаеробів. Найбільш типові факультативні анаероби - дріжджі. Потреба їх в кисні для нормального розвитку далеко не однакова, навіть у грибів, що відносяться до одного і того ж роду. Так, Fusarium oxyporus виживає в анаеробних умовах протягом 13 тижнів, тоді як F. eumartii гине менше ніж через три тижні.
Вологість середовища. Більшість грибів для свого зростання потребує порівняно високій вологості. Так, їстівні гриби зазвичай з'являються в дощову теплу погоду. Розвиток пліснявих грибів також можливо лише на субстратах, що відрізняються підвищеною вологістю. Дереворазрушающие гриби найбільшу швидкість росту мають при абсолютній вологості деревини 30-80%, хоча мінімальна і максимальна вологість, при якій вони розвиваються, знаходиться в межах від 20 до 150%.
Вологість деревини може служити фактором, що обмежує зростання і руйнівну діяльність ряду грибів. Цю особливість використовують, зокрема, для запобігання деревини від ураження грибами, насичуючи її вологою методом додаткового зволоження або дощування. Цей прийом застосовують для підвищення збереження свежесрубленной деревини. Методика його детально розроблена А. Т. Вакін.
Органи статевого розмноження зазвичай розвиваються в повітряному середовищі, і для них більш сприятлива дещо більша сухість в порівнянні з оптимумом для росту міцелію. Показово також, що у одних грибів, переважно цвілевих, навіть невелике зниження вологості повітря затримує утворення спор, в той час як у інших, наприклад мучністоросяних Erysiphales, число утворюються конідій значно збільшується.
Спори грибів відрізняються високою стійкістю до несприятливих факторів і в умовах низької вологості довго зберігають життєдіяльність.
Існують гриби, переважно з гастероміцетов, що пристосувалися до життя в посушливих пустельних умовах (види пологів Simblum, Podaxis і ін.). Вони виносять повне зневоднення, а в період дощів відновлюють свою життєдіяльність. Розвиток грибів на розчинах поживних речовин може стримуватися осмотичним тиском навколишнього середовища, т. Е. Вода в цьому випадку стає недоступною, підвищення концентрації осмотично діяльних речовин викликає плазмоліз клітин і повністю зупиняє їх зростання. Разом з тим величина переноситься клітинами осмотичного тиску неоднакова і визначається як специфічними особливостями грибів, так і природою знаходяться в розчині речовин. За даними В. Ліллі та Г. Барнет, гриб Rhizopus nigricans переносить осмотичнийтиск глюкози в 63,2-10 2 кПа (63,2 атм), сахарози- 42,1-102 кПа (42,1 атм), KNО 3 - 27,5-10 2 кПа (27,5 атм). Сисна сила клітин міцелію різних штамів грибів Trichoderma знаходиться в межах (45-48) 10 2 кПа (45-48 атм). Високою стійкістю до осмотичного тиску розчину відрізняються фітопатогенні гриби, сосущая сила клітин яких значно вище, ніж клітин рослини-господаря.
Забруднення повітря. Зміст в повітрі промислових відходів в підвищених концентраціях чинить негативний вплив на ростові процеси грибів. Так, обробка сірчистим газом 5-денний культури Heterobasidion annosum в концентрації 134 мг / л знижувала ріст міцелію в залежності від штаму на 75-95%. При введенні NO x в концентрації 66,7 мг / л у 7-денну культуру зростання міцелію сповільнювався на 45%, а у деяких штамів взагалі припинявся. Приблизно аналогічна залежність виявлена і в накопиченні біомаси під впливом SО 2 і NO x. У дослідах з впливом важких металів знайдено, що найбільшою токсичністю до Н. annosum володіють солі свинцю і нікелю, в меншій мірі - солі міді і марганцю, а солі цинку в випробовуваних концентраціях ріст кореневої губки НЕ інгібували.
Визначення вмісту фтору в плодових тілах їстівних грибів порядку Agaricales показали, що вміст фтору залежить від виду гриба, місця і року збору і може досягати 1 мг на 100 г сухої речовини, т. Е. Свідчить про вплив навколишнього середовища на вміст фтору в плодових тілах . Як правило, концентрація мінеральних елементів в капелюшках вище, ніж в ніжках.
Доведено, що плодові тіла їстівних грибів здатні акумулювати кадмій і ртуть, мічені радіоактивними ізотопами. Так, у Pleurotus flabella-tus виявлено 75% внесеного в субстрат кадмію і 38,5% ртуті, в той час як у Agaricus bisporus відповідно 1,27% і 8,42%. Таким чином, перенесення металів в плодові тіла у різних видів відбувається з неоднаковою інтенсивністю.
література
1. Жданова М.М., Василевська А.І. Екстремальна екологія грибів. - К., 1982
2. биосинтетической діяльність вищих грибів / О.М. Шивріна, О.П. Мезковская, М.М. Фаліна і ін. - Л., 1969
3. Беккер 3.Е. Фізіологія грибів та їх практичне використання. - М., 2003
4. Денисова Н.І. природа і біологічна рольпротеїназ базидіальних грибів // Мікологія і фітопатологія.- 1984 - Т. 18, №2. -З. 116-121.
Антиметаболіт аргініну, лізину або гістидину, синтезується Act. macrosporus (термофілл). 2. Одиниці біологічної активності. Вираз величин біологічної активності антибіотиків зазвичай виробляють в умовних одиницях, що містяться в 1 мл розчину (од / мл) або в 1 мг препарату (од / мг). За одиницю антибіотичної активності приймають мінімальну кількість антибіотика, здатне придушити ...
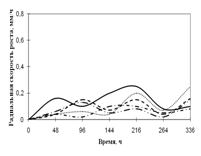
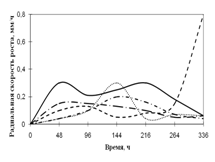
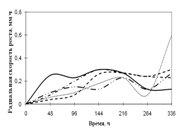
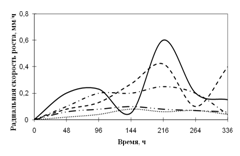
Переглянутих. Спостереження проводять на 1, 2, 3, 7, 15-ту добу. Скло мікроскопіровать, враховуючи проростання суперечка (%), довжину проростків (в мкм) (Кураков, 2001). РОЗДІЛ 3. ДОСЛІДЖЕННЯ ЗРОСТАННЯ мікроміцетів в різних субстратах Об'єктами дослідження стали 10 штамів колекційних мікроскоскопіческіх грибів пологів Aspergillus: A. niger, A. ustus, A. terreus, A. flavus, A. fumigatus, а також ...
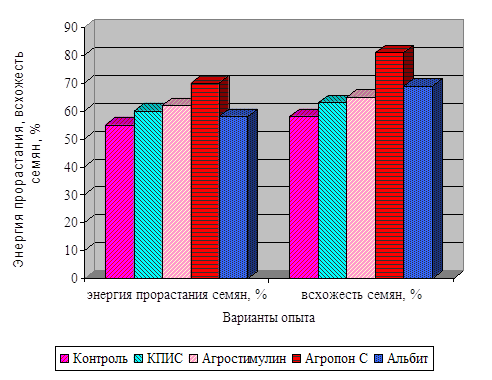
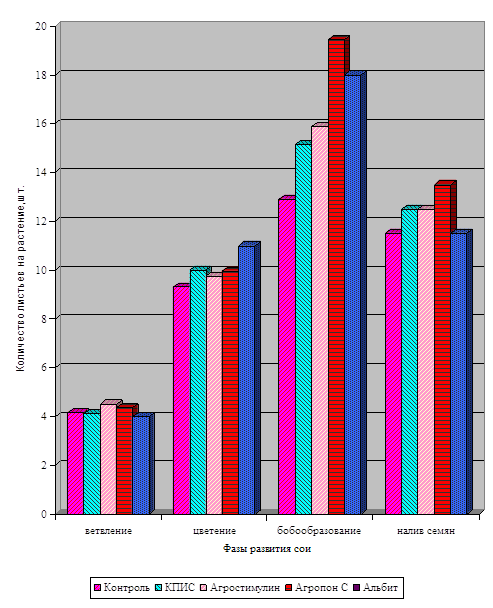
![]()
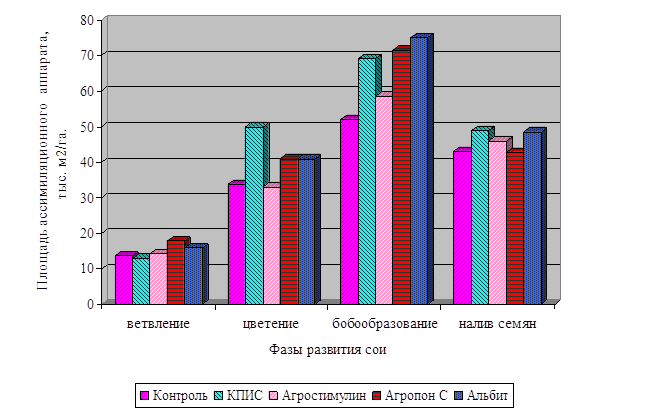
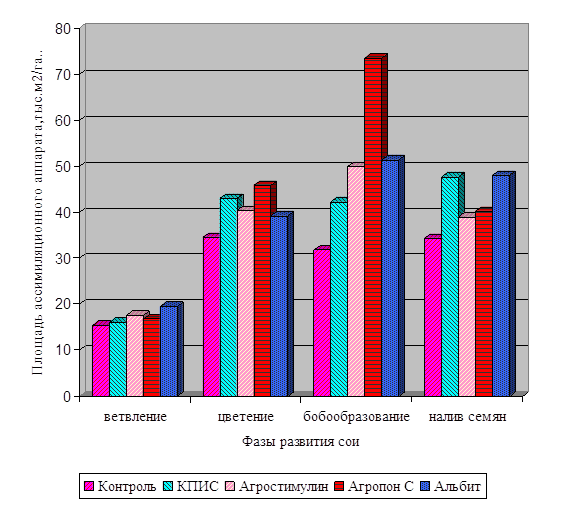
І хвороб. Але ці методи в основному негативно позначаються на якості продукції і на навколишнє середовище. У наших дослідженнях з вивчення впливу регуляторів росту рослин на фізіолого-біохімічні показники та врожайність сої ми застосовували тільки екологічно безпечні препарати. Це продукти життєдіяльності мікроорганізмів і ідентичні за своїм походженням природних сполук, ...
Препаратів. Масштаби цих робіт швидко зростали і продовжують розширюватися в даний час. Синтезовані багато тисяч найрізноманітніших хімічних сполук, багато з яких виявилися високоефективними регуляторами росту та інших життєвих процесів рослин. У нашій країні роботи з вивчення дії і застосування синтетичних регуляторів росту стали швидко розвиватися з 1936 р На ...
сторінка 1
Клітини грибів, як правило, грамположительни. Забарвлення по Граму допомагає виявити супутні бактеріальні клітини і виявити капсулу у криптококків. Тушевие (негативна) забарвлення по Буррі дозволяє виявити інкапсульовані клітини Cryptococcus neoformans в препаратах ліквору при криптококовому менінгіті. Забарвлення за Романовським-Гімзою використовується для виявлення в мокротинні трофозоітов Pneumocystis carinii, у крові або кістковому мозку (як і забарвлення по Райту) - дріжджову форму збудника гистоплазмоза. Для виявлення нокардій та інших кислотостійких грибів застосовують метод забарвлення за Цілем - Нільсеном або модифікований метод Хенкса.
Клітини грибів (гіфи) покриті оболонкою, по складу відрізнялася від бактеріальних і рослинних оболонок; компонентами грибних оболонок є полісахариди і хітин. Належність до певного класу за В. І. Білай до деякої міри зумовлює хімічний складклітинних оболонок. Так, в класі фікоміцетів (підклас зігом-цетов) у багатьох видів оболонка складається з целюлозно-Хіті-нового комплексу; у деяких видів з класів сумчастих і недосконалих - хітин-глюкановий, у дріжджів - Манна-глюкановий. Отже, до складу клітинних оболонок грибів входять в якості основних компонентів: глюкани, хітин, білки і жири. Хітин - речовина, властиве тваринному світу. З хітину складаються жорсткі покриви комах. Для грибних оболонок целюлоза не характерна, але в оболонках, наприклад фітофтори, вона виявлена.
Клітини Грибов і водоростей за своєю організацією схожі на клітини вищих рослин. До складу оболонки входить целюлоза. Протоплазма являє собою складне колоїдне освіту з різко вираженим поверхневий натяг. У цій колоїдної системі безперервної фазою є вода, а дисперсною фазою - ліпопротеі-нові сполуки. У протоплазмі одноклітинних грибних організмів-дріжджів - легко виявляються вакуолі, що представляють собою порожнини, заповнені клітинним соком. При розподілі вакуолі дочірньої клітини утворюються шляхом отпочкови-вання від вакуолі материнської клітини. У протоплазмі є також дрібні гранули - - рибосоми (мікросоми), розміри яких становлять 200 ммк, виявити їх можна лише методом електронної мікроскопії.
Клітка грибів в більшості покрита твердою оболонкою - клітинною стінкою.
Клітини грибів і водоростей за своєю організацією схожі на клітини вищих рослин. До складу оболонки входить целюлоза: Протоплазма являє собою складне колоїдне освіту з різко вираженим поверхневий натяг. У цій колоїдної системі безперервної фазою є вода, а дисперсною фазою - ліпопротеі-нові сполуки. У протоплазмі одноклітинних грибних організмів - - дріжджів - легко виявляються вакуолі, що представляють собою порожнини, заповнені клітинним соком. При розподілі вакуолі дочірньої клітини утворюються шляхом отпочкови-вання від вакуолі материнської клітини. У протоплазмі є також дрібні гранули - рибосоми (мікросоми), розміри яких становлять 200 ммк, виявити їх можна лише методом електронної мікроскопії.
Клітини грибів за своєю будовою істотно не відрізняються від бактеріальних і складаються з тих же елементів.
Поступаючи в клітини грибів, деякі фунгіциди здатні впливати на внутріклетрчние мембрани. Передбачається, що дане з'єднання ннгібірует ліпідний метаболізм, порушуючи синтез таких компонентів клітинних мембран, Як жирні кислоти і стероїди.
Пригнічує поділ клітин грибів.
Ядро в клітинах грибів і водоростей є чітко оформлене освіту, оточене мембраною. На деяких стадіях ділення клітин мембрана зникає. При діленні клітин в ядрах з'являються специфічні освіти - хромосоми.