Клітини основної тканини мають. Покривні тканини. Первинні і вторинні
покривні тканини
Покривні тканини рослин. Покривні тканини. Основні функції - захист молодих органів від висихання, механічний захист і газообмін. Розрізняють: епідерму, перидерму і кірку. 1. Епідерма, первинна покривна тканина. Утворена одним шаром клітин, що покривають всі молоді органи рослин. Покривна тканина зони всмоктування коренів називається епіблемой (ризодерма).
Покривні тканини є прикордонними. Вони розташовуються на поверхні органів рослин, що обумовлює особливості їх будови і різноманіття виконуваних функцій.
1. Покривні тканини забезпечують зв'язок рослин з навколишнім середовищем. При цьому вони є не просто пасивним бар'єром, а й активними учасниками взаємодій. Наприклад, клітини епідерми беруть участь у визначенні сумісності пилку і рильця маточки; в розвитку процесів, пов'язаних з імунітетом рослин, реакцією на зміну температури і вологості повітря і грунту.
2. Покривні тканини здійснюють транспирацию і газообмін і їх регулювання.
3. Покривні тканини беруть участь у виділенні продуктів метаболізму.
4. Покривні тканини захищають рослини від ураження хвороботворними вірусами, бактеріями, грибами; від поїдання тваринами.
5. Покривні тканини оберігають рослини від добових і сезонних перепадів температури.
З урахуванням походження, місця розташування, особливостей клітинної будовиі виконуваних функцій серед покривних тканин виділяють епідерму, епіблему, веламен, перидерму і кірку. На думку ряду вчених сюди саме можна сказати і насіннєва шкірка.
Первинні покривні тканини. Епідерміс. Епіблема. веламен
Епідерма (епідерміс) є первинною покривною тканиною, характерною для всіх вищих рослин. У покритонасінних вона розташовується на поверхні листя, стебел і їх видозмін, а також на репродуктивних органах - квітках і плодах. Крім того, епідермальні клітини можуть вистилати внутрішню поверхню ендокарпій - внутрішнього шару плодової оболонки. Епідерма характерна для трав'янистих рослин. У дерев і чагарників вона розвивається на молодих частинах пагонів, а в міру їх старіння заміщається перидермой.
Епідерма виникає з протодерми, породжується тунікою конуса наростання пагона. Це складна тканина, до складу якої входять основні і побічні клітини, які замикають клітини продихів, а також трихоми - вирости епідерми. Клітини епідерми розташовуються одношарово. Однак у деяких тропічних рослин (пальми, орхідеї та ін.) Епідерміс буває багатошарової.
Основні клітини епідерми стебел, черешків листя, а в однодольних і листових пластинок, мають подовжену форму, витягнуту вздовж осі органу. Наприклад, в стеблах злаків довжина клітин епідерми досягає 300 мкм. У листових пластинках більшості дводольних, на квітках і плодах епідермальні клітини мають Паренхімні і складчасту форму. Основні клітини епідерми містять розвинені вакуолі, але позбавлені хлоропластів і не беруть участі у фотосинтезі. Їх клітинні оболонки мають ряд особливостей. Внутрішня оболонка тонка, гладка. Антиклинальная, тобто поперечна, часто буває звивистою, що збільшує площу зіткнення суміжних клітин епідерми, а отже - силу їх зчеплення. Зовнішня оболонка найбільш міцна. Вона утворена більш товстим шаром целюлози. Хімічний склад оболонок змінюється в ході онтогенезу. У хвощів, осок і злаків оболонки часто просочуються солями кальцію, у вівса - кремнеземом, у цукрового очерету - суберином. Рідше, у бамбуковідних мятлікові, спостерігається одревеснение оболонок. У цитоплазмі епідермальних клітин можуть утворюватися кристали: Рафіду (виноград, недоторка), друзи (буряк), цістоліти (фікус), рідше - кристалічний пісок (картопля).
Протопласт епідерми активно виробляє і виділяє на зовнішню поверхню високомолекулярні органічні сполуки, які в сукупності утворюють кутикулу. У складі катікули перший шар утворюють розгалужені молекули пектину, потім розташовуються грудочки рослинного воску, кутин і епікутікулярний віск. Залежно від щільності складення і скульптування поверхневого шару, кутикула може бути щільною і блискучою, як у листя фікуса, або пухкої і матовою, як у стебел жита, плодів сливи або яблуні. Кутикула знижує інтенсивність транспірації, відображає промені сонця і захищає лист від перегріву, зменшує можливість проникнення патогенів в рослину.
Серед основних клітин епідерми на верхній стороні листа мятлікові зустрічаються великі моторні клітини з великими вакуолями. У спекотну пору вони забезпечують згортання листа в трубку (кукурудза) або складання листової пластинки (ковила), що захищає рослину від надмірного випаровування вологи.
На основних клітинах епідерми часто утворюються вирости, або трихоми, які бувають по виконуваних функцій кроющими і залозистими, а за структурою - волосками і емергенцамі. Кроющие волоски заповнені повітрям і мають білясту забарвлення. Вони добре відображають прямі сонячні промені і зменшують випаровування води. Залізисті волоски - це живі клітини, що виробляють і виділяють ефірні масла і інші кінцеві продукти метаболізму. Волоски утворюються з клітин епідерми. Вони бувають одноклітинними і багатоклітинними, простими і гіллястими. Одноклітинні прості волоски характерні для термопсису і підмаренника, одноклітинні розгалужені - для пастушої сумки і матіоли. Багатоклітинні прості зустрічаються у пеларгонії, багатоклітинні гіллясті - у лаванди, а зірчасті - у лоха і обліпихи.
Емергенци - це поверхневі структури, утворені клітинами епідерми і більш глибоко розташованих тканин. До них відносяться шипи на плодах у селерових, шипи на стеблах малини і ожини, пекучі волоски листя і стебел кропиви. Вони захищають рослини від поїдання.
Замикають клітини продихів розташовуються між основними клітинами епідерми. Вони є активними учасниками транспірації і газообміну. Замикають клітини мають невеликі розміри, містять хлоропласти і попарно розташовуються в епідермі, що не зростися між собою. Між ними знаходиться устьичная щілину, а під ними - подустьічная повітряна порожнина.
У однодольних замикають клітини мають гантелевідной форму. Вони звужені в середній частині і розширені по краях. У розширеній частині клітинні оболонки тонкі і розтяжні, а в середині - потовщені і нерозтяжні. У дводольних замикають клітини мають, при перегляді зверху, бобовидную форму. У них потовщені і нерозтяжна ділянки оболонки, звернені до устьичной щілини. Зовнішні оболонки цих клітин мають вирости, які підносяться над устьичной щілиною і утворюють невелику порожнину - передній дворик. Якщо такі ж вирости виникають нижче устьичной щілини, то вони утворюють задній, або внутрішній, дворик, який безпосередньо пов'язаний з подустьічной повітряної порожниною. У сукупності замикають клітини, устьичная щілину, передній і задній дворик, а також подустьічная повітряна порожнина утворюють продихи, що регулює транспірацію і газообмін. Число продихів досить велике. У культурних рослин на 1 мм 2 листкової поверхні розміщується від 100 до 700 продихів.
Робота продихів зводиться до регулювання ширини устьичной щілини за допомогою складних фізіологічних процесів. Устячка можуть відкриватися в різний часдіб. Це залежить від фізіологічного стану рослин, хімічного складуі концентрації клітинного соку, забезпеченості клітин водою і вуглекислим газом, Освітленості листа, температури повітря і сили вітру.
Ранкове відкривання продихів можна схематично представити наступним чином. Під впливом світла в хлоропластах замикаючих клітин проходить фотосинтез. В результаті концентрація їх клітинного соку збільшується, що призводить до вступу води з сусідніх, побічних, клітин і зростанню тургорного тиску. Під дією тургорного тиску тонкі ділянки оболонок замикаючих клітин розтягуються в сторони, протилежні від устьичной щілини, і товсті ділянки оболонки відходять один від одного. Устьіце відкривається. При зниженні тургорного тиску в замикаючих клітинах продихи закриваються. Детально робота устьиц і процес транспірації вивчаються в курсі фізіології рослин.
Побічні клітини розташовуються поруч з замикаючими клітинами продихів і беруть участь в регулюванні їх водного режиму. За розмірами вони менше основних і не мають хлоропластів. Розташування побічних клітин є діагностичною ознакою, що використовуються в систематиці рослин. Відомо більше десятка типів устьиц в залежності від взаємного розташування замикаючих і побічних клітин. Наприклад, при аномоцітном типі, характерному для винограду, червоного перцю і інших квіткових рослин, численні побічні клітини не відрізняються за розмірами від основних клітин епідерми. При періцітном, відомому у папоротей - одна побічна клітина оточує продихи. У дводольних часто зустрічається парацітний тип продихи, коли дві невеликі побічні клітини розташовуються паралельно замикаючим. Для однодольних характерний тетрацітний тип, де чотири побічні клітини розташовані навколо продихи так, що площини їх стінок не збігаються з площиною розташування устьичной щілини.
Розташування устьиц на поверхні листа також має систематичне значення. Наприклад, у однодольних вони розташовуються рівними рядами, а у дводольних - розсіяно по всій поверхні листової пластинки. Як правило, продихи розташовуються на нижньому боці аркуша. Це зменшує випаровування води наземними рослинами з плагіотропние, тобто паралельним поверхні Землі розташуванням листя (яблуня, буряк і ін.). Якщо лист тяжіє до вертикального, ортотропними, положенню в просторі, то продихи на ньому можуть розташовуватися з обох сторін, як у кукурудзи або цибулі. У водних рослин, листя яких плавають на поверхні води, продихи знаходяться на верхній стороні, як у типовій для білоруських водойм кубушки жовтої.
Епіблема. Особливим видом первинної покривної тканини є епіблема, яка утворюється меристематические клітинами поверхні кінчика кореня. Клітини епіблеми живі, тонкостінні, з розвиненою вакуолью, розташовуються на поверхні кореня в зоні всмоктування. Одні з них, тріхобласти, утворюють особливі одноклітинні вирости - кореневі волоски, Які активно всмоктують з грунту воду і розчинені в ній речовини. Інші, атріхобласти, кореневих волосків не мають. Життя кореневих волосків обмежена двома - трьома тижнями. Потім вони руйнуються і злущуються. Одночасно, в міру зростання кінчика кореня, на ньому утворюються нові кореневі волоски.
Епіблема на коренях однодольних сохранаться протягом усього життя рослини. У дводольних в зв'язку з переходом кореня до вторинного анатомічною будовою епіблема зберігається лише в зоні кореневих волосків, а в зоні проведення вона заміщується перидермой.
Веламен - це первинна покривна тканина повітряних коренів тропічних епіфітів і лазять ліан.
Епіфіти виростають в щілинах гілок, заповнених гумусом, і у підстав листя великих дерев, використовуючи їх лише як опору або субстрат, наприклад, теніофіллум і фаленопсис з сімейства орхідних. Вони поселяються також на скелях. Лазять ліани, до яких відносяться культурні види - ваніль плосколистная (сем. Орхідних) і монстера делікатесна (сем.Ароідние), як субстрат використовують грунт. У них утворюються довгі стебла, що прикріплюються до опорних деревам за допомогою повітряних коренів, що дозволяє виносити листя і квітки в верхній ярус, до світла. У багатьох з них, як у монстери, на стеблі утворюються живлять повітряні коріння, що ростуть вниз. Досягнувши грунту, вони проникають в неї, розгалужуються і всмоктують воду і мінеральні речовини.
Веламен утворюється з протодерми і складається з декількох шарів мертвих клітин, оболонки яких мають сітчасті або спіральні потовщення, що надають їм міцність. Завдяки наявності в оболонках численних пір і наскрізних отворів, веламен як губка капілярним шляхом всмоктує дощову воду, а також конденсується атмосферну вологу. Таким чином, веламен виконує захисну роль і регулює водний режим рослин.
Вторинний покривний комплекс - перидерма
Перидерма є вторинним покривним комплексом, який характерний для голонасінних і дводольних покритонасінних. Вона утворюється на коренях і кореневищах, бульбах і стеблах, на листових рубцях і криють чешуях зимуючих бруньок, рідше - на плодах. Раневая перидерма формується на травмованих ділянках органів рослин.
Перидерма породжується клітинами коркового камбію, іменованого феллогеном. Феллоген має всі ознаки освітньої тканини. Це невеликі прямокутні, злегка потовщені в радіальному напрямку, щільно складені клітини з тонкими оболонками. Вони мають густу в'язку цитоплазму і численні органели, пов'язані з процесами синтезу, що забезпечують поділ ядра і цитоплазми.
Феллоген утворюється, як правило, в перший рік життя органу, наприклад, на пагонах яблуні або смородини. Однак у тиса ягідного він з'являється на 2 - 3 рік, а у клена американського - на 3 - 4 рік. Феллоген закладається в поверхневих шарах рослин. Наприклад, у троянди він виникає в епідермі, у малини - в первинній корі, у спіреї - в періцікле, у пузиреплодник - у вторинній флоеме. При формуванні вторинного анатомічної будови кореня феллоген утворюється з клітин перициклу, а при утворенні кірки повторні шари феллогена виникають у вторинній корі.
Мітотичний поділ клітин феллогена призводить до утворення феллеми і феллодерми. Феллема відкладається назовні від феллогена. Її клітини розташовуються багатошарово, рівними рядами, одні над іншими. У різних групрослин клітини феллеми відрізняються товщиною і видозмінами оболонок, наявністю кристалів оксалату кальцію. При просочуванні оболонок феллеми суберином утворюється пробка. Можливо також утворення феллоіда, тобто тканини, яка з'являється при просочуванні оболонок феллеми лігніном.
Мертві клітини пробки мають щільне складання, вони заповнені повітрям, або загусла протоплазми темного кольору. Пробка непроникна для води і має низьку теплопровідність, тому ефективно захищає рослини від втрати води, перепадів температури, хвороботворних мікроорганізмів. Розвиток пробки має прямий вплив на збереження бульб картоплі і коренеплодів в період зростання і зберігання. Пробка коркового дуба має велике значення у виробництві якісних вин. Феллодерми, утворена феллогеном, є живою паренхимной тканиною. Її клітини містять хлоропласти і здатні накопичувати крохмаль. Пробка в сукупності з феллогеном і феллодерми становить вторинний покривний комплекс - перидерму. В окремих місцях перідерми утворюються чечевички, які служать для газообміну і регулювання водного режиму рослин. Чечевички - це отвори в пробці, заповнені виконує тканиною, що складається їх пухко складених, слабо опробковевшей паренхімних клітин. Під чечевички в феллогене є невеликі межклетники, що полегшують газообмін живих клітин феллодерми. В кінці літа під виконує тканиною чечевички феллоген виробляє шар дрібних, щільно складених клітин замикає шару, який значно зменшує газообмін і випаровування води через чечевички в період сезонного спокою рослин. Навесні замикає шар руйнується під тиском знову утворених клітин пробки і чечевички продовжують виконувати свої функції. Тривалість збереження перідерми залежить від видових особливостей рослин. Найбільш коротка вона у трав'янистих рослин. У садових чагарників (смородина, аґрус) перидерма зберігається все життя, у плодових зерняткових порід (яблуня, груша) - протягом 5 - 10 років. Бук, граб, черемха, ліщина протягом усього життя залишаються гладкоствольною.
кірка
У більшості дерев на зміну перидерма утворюється кірка або рітідом. Кірка виникає з поверхневих шарів кори в результаті закладення повторних перидерм. Феллоген цих перидерм формується з паренхіми вторинної флоеми. При цьому живі паренхімні клітини кори, ізольовані шарами пробки, відмирають і в сукупності з іншими тканинами кори і шарами повторної пробки утворюють кірку. Якщо повторні перідерми утворюються у вигляді неправильних дуг, які своїми кінцями стикаються з раніше виниклими перидерма, то утворюється тріщинувата, або луската, кірка (дуб, клен та ін.). При закладенні повторної перідерми суцільним кільцем, як у винограду, кірка буде кольчатой. При потовщенні стовбура мертві тканини кірки розтріскуються в окремих місцях, однак тріщини не доходять до живих тканин кори. Темпи наростання кірки в товщину невеликі, так як її зовнішні шари поступово злущуються. Кірка виконує захисну функцію, оберігаючи стовбури дерев від механічних пошкоджень, перепадів температури, проникнення патогенів. Однак в її тріщинах можуть зберігатися зимуючі стадії шкідливих комах, спори хвороботворних грибів і бактерій. Це обумовлює необхідність постійного догляду за поверхнею стовбурів деревних садових порід.
насіннєва шкірка
Особливим типом покривної тканини є насіннєва шкірка або насіннєва оболонка, яка утворюється з покривів (інтегументов) семязачатка і є обов'язковою частиною насіння. Вона складається з декількох шарів щільно складених клітин і виконує захисну роль, оберігаючи зародок насіння і запасають тканини від пересихання, механічних пошкоджень і хвороботворних мікроорганізмів. Ослизнение насіннєвих оболонок, як у бобових, льону та інших рослин, сприяє всмоктуванню вологи з ґрунту. Крім того, міцна шкірка може захищати насіння від передчасного проростання, що в агрономічної практиці називається твердокам'яну. У зв'язку з цим насіння деяких культур (конюшина, галега і ін.) Скарифицируют перед посівом, тобто руйнують їх насіннєві оболонки або шляхом механічного перетирання з піском та іншими матеріалами, або впливом сірчаної або інших кислот. Вирости насінної оболонки у вигляді волосків опушення, як у верб і бавовнику, забезпечують поширення насіння вітром. Товщина шкірки часто пов'язана з розвитком плодової оболонки. У рослин з міцним не розкриваються околоплодником вона, як правило, більш тонка, наприклад у горіхоплідних. Важливими структурними утвореннями на поверхні шкірки є микропиле і рубчик. Мікропіле - це невеликий отвір, через яке починається проникнення води при проростання насіння. Рубчик є місцем прикріплення насіння до семяножка.
покривні тканини
покривні тканини- зовнішні тканини рослини.
Вони оберігають органи рослини від висихання, від температурних впливів, механічних пошкоджень та інших несприятливих впливів навколишнього середовища. При змінах віку органів і їх функцій, покривні тканини закономірно змінюють одна одну. За походженням розрізняють первинні (епідерміс, ризодерма, веламен), вторинні (перидерма) і третинні (кірка або рітідом) покривні тканини.
Первинні покривні тканини Епідерма утворюється з поверхневого шару апікальної меристеми - протодерми. Вона покриває листя, плоди, частини квітки і молоді стебла. Крім захисної функції, епідерміс регулює процеси транспірації і газообміну, бере участь в синтезі різних речовинта ін. До складу епідерми входить кілька морфологічно різних клітин: основні клітини епідерми, що замикають і побічні клітини продихів, трихоми (вирости епідерми). Клітини епідерми живі, мають ядра, лейкопласт, вакуолі, хлоропласти (тільки в замикаючих клітинах продихів). Епідерма у більшості рослин одношарова, рідше багатошарова. Клітини первинної покривної тканини щільно прилягають один до одного, і не мають межклетников. Із зовнішнього боку вся епідерміс покрита суцільним шаром кутикули (переривається тільки над устьічнимі щілинами).
Ризодерма (епіблема) утворена апикальной меристемою кореня. Вона покриває молоді кореневі закінчення і саме через ризодерма відбувається поглинання води і мінеральних солей з грунту. Крім того, вона взаємодіє з мікроорганізмами грунту, з кореня в грунт виділяються речовини, що допомагають грунтовому харчуванню. Клітини ризодерма мають дуже тонкі оболонки. У первинної покривної тканини кореня немає кутикули, внаслідок чого ці клітини мають оболонки легко проникні для води. На невеликій відстані від кінчика кореня утворюються кореневі волоски - вирости ризодерма.
Веламен, як і ризодерма, відбувається з поверхневого шару апікальної меристеми кореня. Ця своєрідна тканина покриває коріння епіфітів і деяких інших рослин, пристосованих до життя на періодично пересихаючих ґрунтах (аспідістра, аспарагус, алое, кливия). Веламен від ризодерма відрізняється багатошаровістю. Протопласт Велам відмирає і тому всмоктує воду не осмотическим, а капілярним шляхом.
Вторинна покривна тканина Перидерма виникає при закладенні феллогена в епідермі, субепідермальной шарі (під епідермою) або в більш глибоких шарах первинної кори. Вона заміщає епідерму в тих стеблах і коренях, які розростаються в товщину шляхом вторинного росту. Перидерма складається з трьох основних компонентів: феллогена (корковий камбій), за рахунок якого перидерма тривалий час наростає в товщину, виробляючи до поверхні Фелл (пробку), виконує захисну функцію, а всередину феллодерми (підживлює тканину).
Живі тканини, розташовані під пробкою відчувають потребу в газообміні. Для цього в перидерма з самого початку її утворення формуються чечевички - прохідні отвори.
Третинна покривна тканина
Кірка (рітідом) приходить на зміну перідерми. У більшості деревних рослин вона утворюється в результаті багаторазового закладання нових прошарків перідерми у все більш глибокі тканини первинної кори. Живі клітини, укладені між цими прошарками відмирають. Таким чином, кірка складається з чергуються шарів пробки і укладених між ними відмерлих інших тканин первинної кори.
| біологічні тканини | |
|---|---|
| клітка | |
| тварини | Епітеліальна Сполучна (кісткова, хрящова, жирова, кров і лімфа) Нервова М'язова |
| рослини | Освітня (меристема) покривнаМеханічна Адсорбційна Асиміляційна Провідна Секреторна аеренхіма |
| Див. також | Гістологія Міжклітинна речовина |
| орган | |
Wikimedia Foundation. 2010 року.
Дивитися що таке "Покривні тканини" в інших словниках:
Тканини рослин, розташовані на кордоні з зовн. середовищем. Складаються з щільно зімкнутих клітин. Первинна покривна тканина (епідерміс, або епідерміс) розвивається на листках і молодих стеблах. Товсті зовніш. стінки її клітин покриті кутикулою, ... ...
Рослин зовнішні тканини, що захищають рослини від зовнішніх несприятливих впливів і регулюють повідомлення внутрішніх тканин із зовнішнім середовищем (поглинання і виділення речовин, газообмін, транспірація). Первинна покривна тканина епідерміс, або ... ... Великий Енциклопедичний словник
Рослин, зовнішні тканини, що захищають рослини від зовнішніх несприятливих впливів і регулюють повідомлення внутрішніх тканин із зовнішнім середовищем (поглинання і виділення речовин, газообмін, транспірація). Первинна покривна тканина епідерміс, або ... ... енциклопедичний словник
покривні тканини- тканини, що розташовуються на поверхні органів рослин і захищають їх від втрати води і впливу несприятливих умов зовнішнього середовища. Див. Також епідерміс; перидерма; кірка ... Анатомія і морфологія рослин
Рослин, зовнішні тканини, що захищають рослини від зовнішніх несприятливих впливів і виконують функції поглинання і виділення; через них здійснюється газообмін між рослинним організмомі зовнішнім середовищем. Розрізняють первинні і ... ... Велика Радянська Енциклопедія
Рослин, зовніш. тканини, що захищають р ня від зовн. несприятливих впливів і регулюють повідомлення внутр. тканин з зовн. середовищем (поглинання і виділення в в, газообмін, транспірація). Первинна П. т. Епідерміс, або шкірка, вторинна перидерма ... Природознавство. енциклопедичний словник
покривних тканин- комплекси зімкнутих клітин, що покривають органи рослин, службовці, головним чином, для захисту їх oт несприятливих зовнішніх умов. У листя і стебел вони несуть захисні функції, Біля коріння, крім того, служать для поглинання води і розчинів ... ... Словник ботанічних термінів
ТКАНИНИ РОСЛИН- групи або комплекси клітин, пов'язані спільністю будови, походження, функцій і місця розташування. Відповідно до цього виділяють освітні, покривні, основні, механічні, провідні та видільні Т. р. Освітні тканини, або ... ... Сільськогосподарський енциклопедичний словник
тканини рослин- тканини рослин, групи або комплекси клітин, пов'язані спільністю будови, походження, функцій і місця розташування. Відповідно до цього виділяють освітні, покривні, основні, механічні, провідні та видільні Т. р. ... ... Сільське господарство. Великий енциклопедичний словник
Системи клітин, подібні за будовою, походженням і функцій, розрізняються за розмірами, формою і розташуванням. До складу тканини входять тканинна рідина (заповнює міжклітинні простору) і знаходяться між клітинами речовини, напр. солі кальцію ... ... Біологічний енциклопедичний словник
Покривні тканини розташовані зовні всіх органів рослин на кордоні з зовнішнім середовищем. Вони складаються з щільно зімкнутих клітин і виконують бар'єрну роль, оберігаючи органи рослин від несприятливих впливів. Ці тканини виникли з виходом рослин на сушу і вельми різноманітні за будовою і функціями.
Покривні тканини надземних органів - епідерміс, пробка - служать для захисту від висихання і для газообміну.
коріння первинного будови, Одягає епіблема, регулююча і забезпечує всмоктування і виділення розчинів. Будова епіблеми ми розглянемо в курсі анатомії кореня.
Коріння вторинної будови у дводольних покриті перидермой і кіркою, а в однодольних - екзодермою.
епідерма (Шкірка). Первинна покривна тканина, що утворюється з протодерми (формується з туніки) конуса наростання пагона на всіх листах, стеблах, а також на квітках, плодах і насінні. Епідерма захищає внутрішні тканини від висихання і пошкоджень, перешкоджає проникненню мікроорганізмів. Одночасно епідерміс забезпечує зв'язок із середовищем - через неї відбуваються транспирация (регульоване випаровування) і газообмін, іноді всмоктування і секреція різних речовин (в тому числі ефірних масел, ферментів і гормонів).
Епідерма - складна тканина, в її склад входять морфологічно різні клітини: основні клітини епідерми; замикають і побічні клітини продихів; трихоми, волоски.
Основні клітини епідермищільно зімкнуті, межклетники відсутні. У епідермі стебел і лінійних листя вони у верхній проекції довгасті, в листі з широкими пластинками - більш-менш ізодіаметріческіе або лопатеві. Бічні стінки, т. Е. Перпендикулярні поверхні, часто звивисті, що підвищує міцність їх зчеплення.
Зовнішні стіни зазвичай товще за інших. Їх внутрішній, найбільш потужний, шар складається з целюлози і пектину. Клітинні стінки можуть просочуватися кремнеземом (ріжучі стебла і листя хвощів, деяких осок і злаків) або містити слизу (епідерміс клейких, легко поширюються насіння льону, айви та ін.).
Із зовнішнього боку вся епідерміс покрита суцільним шаром кутикули. Крім Кутіна в її склад входять вкраплення воску, що ще більше знижує проникність кутикули для води і газів. На поверхні кутикули віск може утворити суцільний наліт, що складається з лусочок, паличок і інших структур. Цей сизий, легко стирається наліт добре помітний на листках капусти або плодах сливи, винограду. Якщо його видалити, то плоди будуть швидше псуватися. Потужність кутикули і її склад багато в чому визначають хімічну стійкість і проникність епідерми. В умовах посушливого клімату у рослин розвивається більш товста кутикула. У рослин, занурених у воду, кутикули немає.
Клітини епідерми мають живий протопласт, зазвичай з добре розвиненою ендоплазматичної мережею і апаратом Гольджі. У деяких рослин (традесканції) в цитоплазмі можна бачити лейкопласт. У водних рослин, папоротей, мешканців тінистих місць, а також і багатьох світлолюбних рослин (герань, соняшник, кактуси) зустрічаються хлоропласти.
З епідерми можуть виникати додаткові бруньки, феллоген; в культурі можна отримати зародишеподобние структури. У деяких, переважно тропічних, рослин епідерміс багатошарова, одна з її функцій - запас води.
устячка- спеціалізовані освіти епідерми, що регулюють газообмін, необхідний для дихання і фотосинтезу, транспірації. Устьіце складається з двох замикаючих клітин, між якими знаходиться устьичная щілину - межклетники. Під нею розташована дихальна подустьічная порожнину. Вона сприяє кращому газообміну між внутрішніми частинами органу і зовнішнім середовищем. Часто до замикаючим клітинам примикають дві або більше побічні клітини, відмінні від основних клітин епідерми. Замикають і побічні клітини є устьічнимі апарат.
Стінки замикаючих клітин потовщені нерівномірно: черевні (звернені до щілини) товщі спинних (примикають до епідерми). Замикають клітини містять хлоропласти з добре розвиненими тилакоїди і численні мітохондрії в активному стані. Устячка можуть знаходиться як у відкритому стані так і в закритому. Механізм відкривання і закривання устьичной щілини заснований на осмотических явищах. При підвищенні тургору тонкі стінки розтягуються, захоплюючи за собою товсті, і устьичная щілину збільшується. При падінні тургору вона закривається, так як замикають клітини приймають початкове положення.
Головна роль у зміні тургору і обсягу замикаючих клітин належить іонів калію. При відкриванні устьиц вони переміщаються із сусідніх клітин в прикінцеві, витрачаючи енергію, яку відшкодовують мітохондрії. Істотне значення має і наявність хлоропластів: в результаті фотосинтезу підвищуються концентрація цукрів і осмотичний тиск. За рахунок всмоктування води обсяг вакуолі істотно збільшується, тургор зростає і продихи відкривається. У темряві при недостатньому обводнюванні устьичная щілину закривається через зниження тургору в замикаючих клітинах.
Епідерма дуже ефективно регулює транспірацію. Якщо продихи відкриті повністю, то транспирация йде з такою ж швидкістю, як якби епідерми не було зовсім. При закритих продихи вона різко знижується.
трихоми- різні за формою, будовою і функції вирости клітин епідерми. Вони мають форму волосків (криють або залізистих), лусочок і ін. Функції більшості типів трихом неясні. Кроющие трихоми можуть бути одноклітинними (у яблуні), багатоклітинними нерозгалуженими (у картоплі) або розгалуженими (у коров'яку), зірчастими (у лоха).
волоскиздатні довго залишатися живими. Але часто протопластів в них відмирають, волоски заповнюються повітрям. Такі волоски захищають рослину від сильної сонячної інсоляції, зайвого випаровування і коливань температури. Багато високогірні рослини (едельвейс) відрізняються сильним опушенням. Деякі мертві волоски, наприклад покривають насіння бавовнику, досягають в довжину 55 мм і широко використовуються в текстильній промисловості. Трихоми захищають рослину від комах - чим густіше опушення, тим рідше комахи використовують його в якості їжі або для відкладання яєць, на гачкуваті трихоми комахи і їх личинки наколюють.
Крім волосків на епідермі утворюються емергенци,у формуванні яких беруть участь більш глибоко розташовані тканини. До них відносять пекучі волоски кропиви, шипи троянди, ожини.
Епідерма функціонує, як правило, один рік, зазвичай до осені її замінює пробка.
пробка(Феллема). Вторинна покривна тканина розвивається з клітин коркового камбію, феллогена. Феллоген - вторинна меристема, він виникає з основної паренхіми, що лежить під епідермою або більш глибоко (смородина, малина), а іноді і в самій епідермі (верба). У більшості дерев і чагарників феллоген закладається в однорічних пагонах вже в середині літа. Клітини феллогена діляться паралельно (║) поверхні органу (Тангентально), відкладаючи назовні клітини феллеми, Всередину - феллодерми.Клітин феллеми утворюється завжди більше, ніж феллодерми. Феллема (покривна тканина, пробка), феллоген ( освітня тканину) І феллодерми (основна тканина, хлорофіллоноснимі паренхіма) - утворюють єдиний покривний комплекс - перидерму.
Клітини пробки з'єднані дуже щільно, без межклетников, їх клітинні стінкиспочатку дуже тонкі, потім товщають. Вторинні клітинні стінки складаються з шарів суберіна і воску, що не пропускають воду і повітря. Обкоркування стінок веде до відмирання протопласта. У міру того як епідерму змінює перидерма, зелений колір пагонів переходить в бурий. «Доспілі» до осені пагони першого року життя, захищені перидермой, здатні до перезімовиванію. Пробка захищає органи рослин від втрати води, проникнення хвороботворних організмів, різких коливань температури, так як має малу теплопровідність.
Газообмін і транспірація в органах, покритих перидермой, відбуваються через чечевички. Чечевички - ділянку перідерми з пухко розташованими клітинами пробки. Чечевички з поверхні виглядає як горбок. За межклетникам цієї виконує тканини циркулюють гази і водяну пару. З настанням холодів феллоген відкладає під виконує тканиною замикає шар з щільно з'єднаних клітин, що перешкоджає випаровуванню. Навесні цей шар під напором знову формуються клітин розривається. У міру потовщення гілок чечевички розтягуються (у берези вони мають вигляд рисочок, у осики - ромбів).
У деревних рослин перидерма утворюється на гілках, стовбурах, корінні і ниркових чешуях, на деяких плодах, в місцях ураження органів; у дводольних трав - на коренях, Гіпокотиль, іноді на кореневищах, бульбах. Найбільш масивна щорічно наростаюча пробка стовбурів коркового дуба, яка використовується в промисловості.
Ступінь сформованості перідерми і її характер необхідно враховувати при виборі способів і режиму зберігання овочів. У перидерма коренеплодів моркви шар пробки тонкий, феллоген тут продукує переважно феллодерми, вона потужніша, ніж феллема. Коренеплоди щоб уникнути пересушування зберігають у піску. Успіх зберігання бульб картоплі залежить багато в чому від сформованості перідерми. З молодих бульб шар пробки легко знімається, так як рвуться живі тонкостінні клітини феллогена. Після того як феллоген диференціюється в постійні тканини, картоплю можна закладати на зберігання.
Лише у деяких деревних (осики, бука, ліщини) перидерма захищає стовбури протягом усього життя, а у більшості - через 10 ... 30 років замінюється кіркою. На стовбурах розвивається кілька перидерм, кожна наступна закладається глибше попередньої. Живі тканини, укладені між шарами пробки, відмирають, і формується покривний комплекс - кірка (рітідом). Кірка складається з декількох шарів пробки і укладених між ними відмерлих тканин. Якщо освіта перидерм відбувається не по всьому колу стовбура, а окремими напівдугою, то кірка формується неправильними шматками. Така кірка називається лускатоїі утворюється у більшості рослин. кільцеподібнакірка формується в тому випадку, якщо кожна знову виникає перидерма оперізує стовбур, періодично відрізаючи циліндричні ділянки кори (наприклад, у винограду).
Кірка не здатна до розтягування, тому при потовщенні стовбура в ній з'являються тріщини. На дні тріщин у внутрішній перидерма є чечевички, що забезпечують газообмін. Кірка надійно оберігає стовбури від механічних пошкоджень, лісових пожеж, різкої зміни температур.
Основні тканини.
Основні тканини складають більшу частину тіла рослини. За походженням основні тканини майже завжди первинні, утворюються з апікальних меристем. Вони складаються з живих паренхімних клітин, сильно варіюють за формою, але в типовому випадку паренхіма основної тканини складається з клітин, у яких довжина трохи більше, ніж ширина, частіше майже ізодіаметріческіх. Клітинна оболонка первинна тонкостінна, з простими порами. Іноді первинна оболонка сильно потовщується, що особливо характерно для запасающей паренхіми насіння деяких рослин (Asparagus, Coffea arabica, хурма Diospyros, фінікова пальма Phoenix dactylifera). Вуглеводи таких оболонок розглядаються як запасні речовини, використовувані зародком під час проростання. Основна паренхіма здатна повертатися до меристематической активності, наприклад при загоєнні ран, утворенні додаткового коріння і втеч.
Основні тканини виконують функції: синтезу, накопичення та використання органічних речовин. Залежно від виконуваної функції розрізняють основну (типову), асиміляційну, запасаючу і воздухоноснуюосновні тканини.
Основна паренхімане має специфічних, строго визначених функцій. Вона розташовується всередині тіла рослини досить великими масивами. Типова основна паренхіма заповнює серцевину стебла, внутрішні шари кори стебла і кореня. Її клітини утворюють вертикальні і горизонтальні тяжі (промені), за якими здійснюється радіальний транспорт речовин. З основної паренхіми можуть виникати вторинні меристеми.
Асиміляційна паренхіма (паренхіма).Головна її функція - фотосинтез. Паренхіма розташована в надземних органах, зазвичай під епідермою. Особливо добре розвинена в листі, менше-в молодих стеблах. Характерно наявність межклетников, що полегшують газообмін. Клітини тонкостінні, в постінному шарі цитоплазми багато хлоропластів. Загальний обсяг їх може досягати 70 ... 80% обсягу протопласта.
Запасающая паренхіма.Служить місцем відкладення надлишкових в даний період поживних речовин. Запасають тканини складаються з живих тонкостінних клітин. У крахмалоносних клітинах пластиди мають досить просте внутрішню будовуі можуть бути класифіковані як амілопласти; вони зустрічаються у багатьох насіння і підземних органів, що запасають. Паренхімні клітини в квітках і плодах часто містять хромопласти. У деяких насіння в паренхімі запасающей тканини або зародка зберігаються твердий білок або жири. У різних органах рослини паренхімні клітини можуть ставати особливо помітними завдяки накопиченню в вакуолях антоціанів або таннинов або внаслідок відкладення в них кристалів тієї чи іншої форми. Клітини запасающей паренхіми можуть мати товсті клітинні стінки (накопичується гемицеллюлоза в насінні фінікової пальми), або накопичують жири.
У цих тканинах накопичуються багато рослинні продукти, що використовуються людиною. У культурних харчових рослин зазвичай гіпертрофовано розвиток запасающей паренхіми. Запасають тканини широко поширені, розвиваються в самих різних органах. Їх можна виявити в бульбах картоплі, коренеплодах буряка, моркви, цибулинах цибулі, зернівках злаків, в насінні соняшнику, рицини, а також в стеблах цукрової тростини, кореневищах, коренях.
У рослин посушливих місць - сукулентів (агави, алое, кактуси) - в клітинах запасающей паренхіми накопичується вода, Також як у рослин засолених місцезростань (солерос). Великі водоносні клітини є в стеблах злаків. У вакуолях водоносних клітин є слизові речовини з високою водоудерживающей здатністю.
Воздухоносной паренхіма (аеренхіма).Виконує вентиляційні, почасти дихальні функції, забезпечуючи тканини киснем. Складається з клітин різної форми (наприклад, зірчастих) і великих межклетников. Добре розвинена в органах рослин, занурених у воду (в квітконіжках латаття, в стеблах пухівки, білокрильника, рдеста, в коренях очерету).
Механічні тканини.
Механічні (опорні) тканини забезпечують міцність рослини, здатність протистояти дії тяжкості власних органів, поривів вітру, дощу, снігу, витоптування тваринами. Механічні тканини мають сильно потовщені клітинні стінки, які навіть після відмирання протопласта продовжують виконувати опорні функцію.
У проростків, в молодих ділянках органів механічних тканин немає, необхідну пружність вони мають завдяки тургору. У міру розвитку органу в ньому з'являються спеціалізовані механічні тканини - колленхіма і склеренхіма.
Коленхіма.
розвивається у дводольних рослин. Коленхіма, яку розглядають як опорну тканину, складається з товстостінних клітин. Клітини коленхіми витягнуті в довжину, містять протопластів з усіма органелами, часто містять хлоропласти, здатні до відновлення меристематической активності. Для коленхіми характерні первинні, потовщені і нелігніфіцірованние ( неодревесневшіе)оболонки клітин.
найбільш характерну особливістьколенхіми становить структура клітинних оболонок. На свіжих зрізах оболонки виглядають товстими і блискучими, потовщення їх часто розподілені нерівномірно. Крім целюлози вони містять велику кількість пектину і геміцелюлози, але в них немає лігніну. Оскільки пектинові речовини гідрофільних, оболонки клітин коленхіми багаті водою, що можна продемонструвати, обробляючи свіжі зрізи коленхіми спиртом. Зневоднює дію спирту викликає помітне стиснення оболонок. На ультраструктурному рівні в оболонках клітин коленхіми різних типів виявляється шаруватість: ламелли з поперечною орієнтацією микрофибрилл чергуються з ламелами, в яких фібрили орієнтовані поздовжньо. В оболонках клітин коленхіми, особливо в тих, які мають досить рівномірну товщину, часто зустрічаються первинні пори. Характер розподілу потовщень клітинної оболонки змінюється в залежності від типу коленхіми.Якщо оболонка потовщена нерівномірно, то найбільшої товщини вона досягає в кутах клітини або на двох її протилежних - зовнішньої і внутрішньої - тангенціальних стінках. Коленхіма з потовщеннями оболонки, локалізованими в кутах клітин, зазвичай називається уголковой Коленхіма, А з потовщеннями на тангенціальних стінках - ламеллярной,або пластинчастої, Коленхіма. З віком тип оболонки може змінюватися внаслідок відкладення в ній додаткових шарів. Так, наприклад, первинний розподіл потовщень оболонки, характерне для уголковой коленхіми, може ставати все більш невиразним в міру того, як просвіт клітини на поперечному зрізі зменшується.
Наявність межклетников не є обов'язковою характеристикою коленхіми. У тому випадку, коли межклетники присутні в уголкового типі коленхіми, до них завжди звернені потовщені стінки клітин. Коленхіма з таким розподілом потовщень оболонки іноді виділяється в особливий тип, званий пухкої (лакунарной) Коленхіма. Якщо колленхіма не утворює межклетников, то в кутах, де з'єднуються один з одним кілька клітин, серединна пластинка потовщується.
Розташування коленхіми в рослині
Для коленхіми дуже характерно периферичний становище. Вона розташовується або безпосередньо під епідермою, або на відстані одного або декількох шарів від неї. У стеблах колленхіма часто утворює суцільний шар, розташований по колу вздовж осі стебла. Вона зустрічається, іноді у формі тяжів, в виступаючих ребрах багатьох трав'янистих стебел і тих дерев'янистих, які ще не вступили в стадію вторинного росту. В черешках колленхіма розподіляється так само, як і в стеблах. У листовій пластинці вона знаходиться в тих жилках, в яких проходять більші провідні пучки, причому іноді вона розташовується по обидва боки жилок, а іноді тільки з однієї, зазвичай нижній, сторони. Коріння рідко містять Коленхіма.
Зв'язок структури з функцією
Коленхіма пристосована, перш за все, для виконання функції опори зростаючих листя і стебел. Функції опорної тканини колленхіма може виконувати тільки в стані тургору. Її оболонки починають товщати на ранніх етапах розвитку втечі, проте утворюється потовщення пластично і здатне до розтягування. Тому воно не перешкоджає подовженню стебла і листа. На більш просунутій стадії розвитку колленхіма як опорна тканину зберігається в тих частинах рослини (багато листя, деякі трав'янисті стебла), Для яких не характерна сильно розвинена склеренхіма. У зв'язку з обговоренням опорної ролі коленхіми цікаво відзначити, що в країнах, що розвиваються частинах рослини, що піддаються механічним навантаженням (дії вітру або вантажу, прикладеного до похилих паросткам), потовщення оболонки починається раніше і стає більш масивним, ніж у рослин, які не піддавалися таким навантаженням.
У старих частинах рослини колленхіма може ставати жорсткою або перетворюватися в склеренхіма шляхом відкладення вторинних здеревілих оболонок. Якщо вона не зазнає цих змін, то її роль як опорної тканини знижується внаслідок розвитку склеренхіми в більш глибоких частинах стебла або паростка. Більш того, в стеблах з вторинним ростом основний опорної тканиною стає ксилема завдяки переважанню в ній клітин з здеревілими вторинними оболонками і достатку довгих перекривають один одного клітин.
Склеренхіма.
Розрізняють первинну і вторинну склеренхіма. Первинна склеренхіма розвинена у всіх вегетативних органах однодольних, рідше дводольних рослин; вторинна - у переважної більшості дводольних. Клітини склеренхіми мають рівномірно потовщені, як правило, здерев'янілих стінки. Їх міцність близька до міцності стали. Порожнина клітки мала, пори прості щілиновидні, нечисленні. Протопласт відмирає рано і опорну функціювиконують мертві клітини. Розрізняють два основних типи склеренхіми: волокна і склереїди.
волокна- сильно витягнуті прозенхімних клітини довжиною від декількох десятих часток міліметра до 1 (кропива) і навіть 4 см (рами). Вони забезпечують міцність органів рослин на розтяг, стиск і вигини. Міцність волокон підвищується завдяки тому, що фібрили целюлози проходять в них гвинтоподібно, змінюючи напрямок у зовнішніх і внутрішніх витках. Кінці клітин частіше загострені (льон), можуть бути гіллястими (конопля), тупими (кропива) і ін.
У багатьох рослин первинні волокна значно довше, ніж вторинні. Так, у конопель первинні волокна досягають 12,7 мм, а вторинні - всього 2,2 мм. для практичного використаннятакі особливості мають істотне значення. Склеренхімние волокна можуть зустрічатися в рослині у вигляді окремих клітин (елементарне волокно) або, з'єднуючись один з одним по довжині, утворюють пучок (технічне волокно). Волокна виділяють за допомогою мочки стебла або механічно. Кращі результати дає мочка, коли паренхімні тканини, що оточують пучки волокон, руйнуються в результаті діяльності бактерій.
Волокна стебел дводольних рослин використовують для виготовлення різних тканин (особливо цінується неодревесневающіе волокна льону) рідше мотузок (пенька, що отримується з конопель).
склереїди- клітини, частіше за все мають Паренхімні форму. Вони можуть розташовуватися в рослині щільними групами або у вигляді поодиноких клітин. Остаточно сформувалися склереїди - це мертві клітини з товстими здеревілими стінками, пронизаним поровимі каналами, нерідко гіллястими. Пори прості. Склереїди мають первинне походження. До них відносяться кам'янисті (брахісклереіди) і гіллясті (астеросклереіди) клітини.
кам'янистіклітини - округлі, зазвичай зустрічаються групами. З них складаються кісточки вишні, сливи, персика і шкаралупа горіха. Вони зустрічаються в соковитих плодах груші, айви, горобини і в коренях деяких рослин. У деяких сортах груш спостерігається раздревесненіе кам'янистих клітин при дозріванні плоду.
гіллястіклітини мають різноманітну форму, грають роль опорних в листі чаю, камелії, маслини, в стеблах водних рослин.
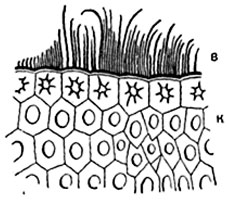
Шкірочка, або епідерміс. Шкірочка, або епідерміс, являє собою первинну тканинулистя і стебел. Шкірочка складається з щільно зімкнутих клітин, що мають в плані у більшості рослин більш-менш звивисті обриси (рис. 86). На поперечних розрізах органу клітини шкірки зазвичай мають чотирикутні або п'ятикутні обриси. Іноді зовнішня і внутрішня стінки злегка випуклі. Клітини шкірки органів, витягнутих у довжину (листових черешків, листових пластинок лінійного і ланцетного обриси, стебел), зазвичай сильно подовжені в напрямку, паралельному поздовжньої осі органу. У деяких рослин, наприклад у багатьох злаків, шкірка складається з клітин декількох типів (рис. 87).
Оболонка епідермальних клітин потовщується зазвичай нерівномірно: в кожній клітині найбільш товста зовнішня стінка, бічні стінки трохи тонше, внутрішня - порівняно тонка. Зустрічаються виключення з цього правила. Бічні і внутрішні стінки клітин зазвичай мають пори (рис. 87), зовнішня ж стінка буває забезпечена ними в дуже рідкісних випадках. Оболонка клітин полягає в основному з целюлози. У багатьох рослин (особливо у злаків, осокових, хвощів) зовнішня стінка шкірки просякнута кремнеземом; у деяких рослин в зовнішній стінці, рідше в бічних (у драцен) і навіть у внутрішніх відкладаються дрібні кристали або зернятка щавлевокислого кальцію.
Поверхня шкірки покрита плівкою - кутикулою (надкожица), що складається з Кутіна. Кутикула дуже тонка або зовсім відсутня на поверхні конусів наростання. Часто кутикула має гладку поверхню, в інших випадках утворює виступи у вигляді горбків, перекладин, прямих або хвилястих ребер і т. Д.
Мал. 86. Ділянки шкірки аркуша жовтця повзучого ( Ranunculus repens):
1 - з верхньої сторони листа; 2 - з нижньої його боку.
Ці виступи складаються без остачі з кутикули або ж є виступами целюлозної стінки, покритими кутикулою.
На поперечних зрізах шкірки іноді видно, що кутикула пронизує і частина товщі стінок. Кутикула сильно заломлює світло і тому добре видно на зрізах. Ізольована кутикула зазвичай представляє безбарвну безструктурну суцільну плівку. У деяких рослин (наприклад, у золотого дерева Aucuba) Кутикула складається з пластинок, кожна з яких належить одній епідермальній клітці; над прикордонними ділянками клітин шкірки кутикули немає або вона дуже тонка.
Потужність кутикули залежить від виду і віку рослини і умов місцеперебування (ріс.88); на поверхні стебел з багаторічною шкіркою (у деяких видів калини, клена) кутикула розривається і підновляється зсередини шляхом утворення нових її нашарувань. Кутикула листя деяких осокових піддається окремненние.
У багатьох рослин в зовнішніх стінках клітин шкірки, іноді в бічних, ще рідше у внутрішніх, утворюються кутикулярного шари- прошарку в целюлозної клітинної оболонці, що містять кутин (рис. 76). Кутикула і кутикулярного шари нерідко просякнуті воском; при підігріванні препаратів в воді під покривним склом він виплавляється в вигляді крапель. У деяких рослин віск покриває кутикулу у вигляді дрібних зерен, розташованих рівномірним шаром (на листках і стеблах багатьох злаків, лілійних, наприклад тюльпанів), тонких паличок, часто вигнутих і на кінці закручених (на стеблах деяких злаків, цукрової тростини, рис. 89) , суцільних кірочок, тонких - 1? товщиною (на листках туй, молодив).
![]()
1 - замикають клітини продихів; 2 - побічні клітини; 3 - клітини епідермісу з численними порами; в порожнині клітин - постінному протоплазма і ядро, в деяких з клітин не змалювати.
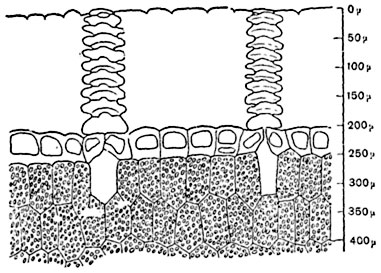
Мал. 88. Частина поперечного розрізу через лістообразние горбок кактуса Ariocarpus retusus; надзвичайно товста кутикула пронизана фігурними ходами над устьицами. Під шкіркою - зелена паренхіма.
Восковий наліт, як і кутикула, знижує транспірацію органів. Віск робить поверхню органів несмачіваемих: з них швидко стікає вода, ніж попереджається капілярний закупорювання водою устьиц і заселення поверхні рослин дрібними епіфітами.
до- шкірка: в- воскові палички.
Тому зрозуміло, чому стебла і листя багатьох підводних рослин позбавлені кутикули.
Клітини шкірки містять тонкий постінному шар протоплазми з ядром та пластидами і велику центральну вакуолю. Зазвичай з пластид тут є лейкопласт.
У багатьох рослин в вакуолях клітин шкірки - нерідко тільки на нижній стороні листа - міститься антоціан.
У вакуолях епідермальних клітин нерідко зустрічаються дубильні речовини, солі щавлевої кислоти у вигляді розчинів (у щавлю, кислиць) або кристалів; рідше тут є алкалоїди, отруйні глюкозиди, гіпс (у каперсів - у вигляді кристалів), кальцієві солі яблучної і винно-кам'яної кислот (у винограду).
Хлоропласти і хромопласти в епідермісі квіткових рослин зустрічаються дуже рідко, тільки у деяких тіньових і водних рослин.
устячка. Для здійснення газообміну між внутрішніми тканинами рослин і зовнішнім середовищем і для транспірації (випаровування води) в шкірці є продихи.
Устьіце складається з двох спеціалізованих замикаючих клітині щелевидная отвори між ними - устьичной щілини. При розгляданні шматочка шкірки, покладеного плазом на предметне скло, продихи зазвичай видно як пара бобовидних або напівмісячних клітин, звернених один до одного увігнутими сторонами (рис. 90-92). Оболонка кожної замикає клітини на внутрішній стороні сильно і нерівномірно потовщена, на протилежній, зовнішньої, сторони - тонка.
На поперечному розрізі, проведеному через середину устьичной щілини перпендикулярно до неї (рис. 91), видно вирости стінок замикаючих клітин продихи, звані відрогами або клювиками.
Щілина продихи має складне обрис: проміжок між зовнішніми відрогами веде в переддень, або передній дворик продихи. Далі йде найбільш вузька частина устьичной щілини - центральна щілина продихи, Що знаходиться між тонкими ділянками внутрішніх стінок. Центральна щілину розширюється далі в задній дворик. Отвір між відрогами нижніх стінок замикаючих клітин
Мал. 90. Шкірочка листа гіацинта ( Hyacinthus orientalis) З устьицами, в плані:
з- замикають клітини, про- щілина продихи.
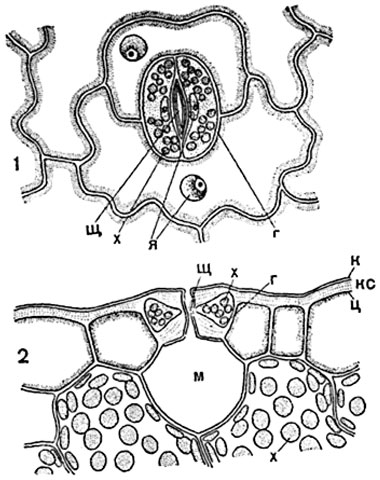
Мал. 91. Устьіце з оточуючими клітинами епідермісу (листя чебрецю Thymus):
1 - в плані; 2 - в розрізі; щ- центральна щілина продихи; а- тонка стінка замикає клітини продихи, звернена до сусідніх клітин шкірки; в замикаючих клітинах видно ядро ( я) З ядерцем і хлорофілові зерна ( x); до- кутикула; м- межклетноє воздухоносной порожнину ( "дихальна порожнина").
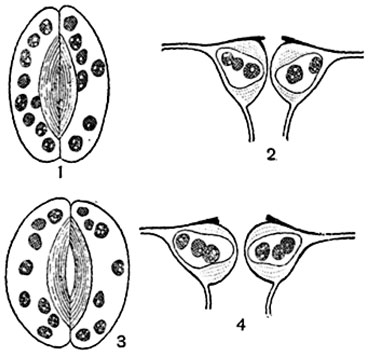
Мал. 92. устьічнимі апарат листа буряка ( Beta vulgaris):
1, 3 - продихи в плані; 2, 4 - в розрізі; 1, 2 - продихи закрито; 3, 4 - продихи відкрито.
повідомляє задній дворик з дихальною порожниною (рис. 91, 2 ; 93).
Повертаючись до продихів в плані (рис. 91, 1 ), Відзначимо, що щілина продихи в оптичних перетинах, паралельних поверхні епідермісу, має форму вузької двоопуклої лінзи; в плані зазвичай видно обрис щілини на рівні зовнішнього отвору і на рівні середини центральної щілини. Зовнішні стіни, а у деяких рослин частково також і внутрішні покриті кутикулою; облямівки двориків іноді складаються суцільно з кутикули. Функціонуючі замикають клітини живі; вони містять постінному шар протоплазми з ядром і з хлорофіловий зернами.
Зазвичай продихи розподіляються на поверхні листа або стебла поодинці і більш-менш рівномірно (рис. 90). У деяких рослин (наприклад, у злаків) вони розташовуються поздовжніми рядами, а в інших (у олеандра, жовтяниці) - групами.
Освіта замикаючих клітин продихів відбувається при нерівномірному розподілі меристематичних клітин епідермісу, в результаті чого виникають ініціальні клітини продихів. У найпростіших випадках ініціальна клітина стає виробляє кліткою продихи.
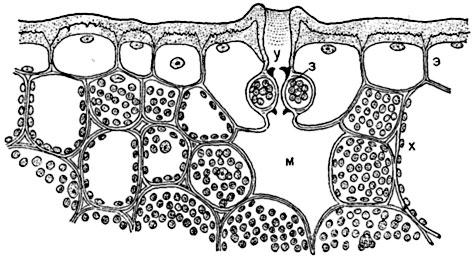
Мал. 93. Частина поперечного розрізу листа Aloe distichaз продихів, зануреним в тканину листа:
е- шкірка з товстими зовнішніми стінками, покритими потужною кутикулою; у- порожнину над продихи; з- замикають клітини з кутикулярними виступами (позначеними чорним кольором); м- воздухоносний межклетники ( "дихальна порожнина"); х- клітини хлорофіллоносной паренхіми.
Виробляє клітина утворює шляхом ділення пару замикаючих клітин продихи; розщеплення перегородки між ними веде до утворення устьичной щілини. У деяких рослин ініціальна клітина утворює шляхом ділення цілі групи клітин - пару замикаючих і дві або кілька побічних клітин продихи; останні відрізняються від інших клітин шкірки меншою величиною і своєрідними обрисами (рис. 94).
Устьічнимі апарат регулює газообмін рослини. Замикають клітини можуть зміщуватися, змінювати обсяг і форму, в результаті чого змінюється і обрис устьичной щілини: вона може бути більш-менш широко відкрита або ж зовсім закрита (рис. 92). При відкриванні та закриванні продихів відбуваються зміни живого вмісту замикаючих клітин. При відкритій щілини продихи протоплазма в них більш в'язка, ніж при закритій. Ядро змінює форму: у кормових бобів при тривало відкритому продихи воно стає веретеновідньш, при закритому - округло-лопатевим, у жоржини - навпаки (рис. 95).
Відкривання і закривання продихів обумовлюється тургорного явищами. Зміни тургору замикаючих клітин можуть бути безпосереднім наслідком подвядания листа або підвищення в ньому вмісту води.
При підвищенні тургору замикаючих клітин обсяг їх клітинної порожнини збільшується, в результаті чого стінки віддаляються одна від одної і щілину відкривається. При зниженні тургору замикаючих клітин відбуваються зміни зворотного роду: стінки зближуються до дотику і щілину продихи замикається.
Зміни тургору в замикаючих клітинах регулюються перетворенням в них крохмалю хлоропластів в цукор, і назад - цукру в крохмаль: при осахаривании крохмалю в замикаючих клітинах підвищується концентрація клітинного соку і збільшується їх сосущая сила. В результаті замикають клітини насмоктують воду (частково за рахунок сусідніх клітин), що викликає зміна їх обсягу та розкриття устьичной щілини (рис. 96). Перетворення цукру замикаючих клітин в крохмаль тягне аналогічним чином закривання щілини продихи.
Непряме участь в закриванні і відкриванні устьиц можуть приймати не тільки клітини, сусідні з замикаючими, а й більш далекі побічні клітини продихів і рядові клітини шкірки. При тривалому дощі все клітини шкірки насичуються водою, тургор їх сильно підвищується і замикають клітини продихів стискаються; в результаті устьічниє щілини пасивно замикаються. Через деякий час після припинення дощу нормальна робота устьиц відновлюється.
Тургорного теорія дії устьиц, висунута Молем в 1856 р, піддалася доповненням з боку ряду вчених, що вказують на можливе значення змін проникності протоплазми замикаючих клітин.
У пристрої устьиц зустрічається велика різноманітність. Форма щілини може бути значно спрощена або ускладнена у порівнянні з вищеописаною, типовою. Найбільш спрощена вона у рослин з листям, плаваючими на воді (у білого латаття): щілину має вигляд воронки, широким кінцем з'єднується з дихальною порожниною, а вузьким переходить в зовнішній отвір; ступінь розкриття щілини продихи залежить від стану цього отвору.
У рослин з листям, розташованими більш-менш горизонтально, особливо ж у дерев і у трав тінистих місць існування, устячка розташовані переважно або виключно на їх нижній стороні.
У багатьох рослин, переважно у приурочених до сухим, сонячним і вітряним місцепроживання, продихи занурені в глиб органу за рахунок сильно кутінізірованних виступів сусідніх клітин (рис. 93). Іноді продихи розташовуються цілими групами в глибині ямок (як у олеандра). У деяких видів австралійських Бенкс ( Banksia) На дні і навколо ямок з устьицами є довгі білі волоски; утворюються, таким чином, притулку з затишшям від вітру і затінення від сонця.
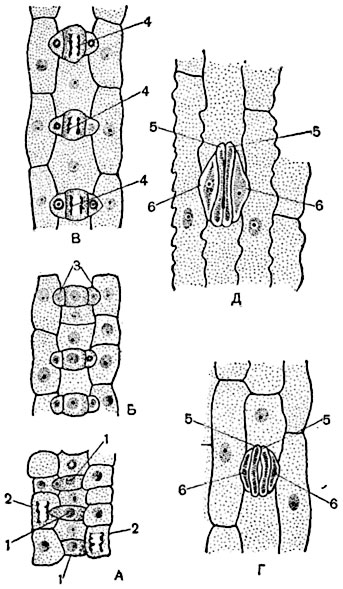
А- три виробляють клітини продихів ( 1, 1, 1 ) Вже відокремилися перегородками від ініціальних клітин; в сусідніх клітках відбувається поділ ядра ( 2, 2 ), Що передує утворенню побічної клітини; Б- за будь-якої з видобувних клітин трьох устьиц утворилися дві побічні клітини ( 3 ); В- в трьох виробляють клітинах продихів відбувається поділ ядра ( 4, 4 ), Що передує утворенню замикаючих клітин; Г- виробляє клітина поділилася на дві клітини, що диференціюються в пару замикаючих клітин продихи ( 5, 5 ), З устьичной щілиною між ними; 6, 6 - побічні клітини; Д- сформований устьічнимі апарат, що складається з двох замикаючих клітин ( 5, 5 ) І двох побічних клітин ( 6, 6 ).
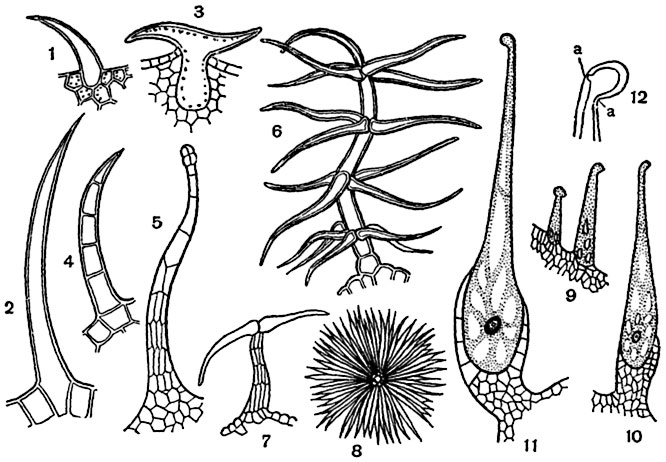
Волоски (трихоми). У більшості вищих рослин деякі або багато з клітин шкірки утворюють вирости, так звані волоски, що мають різну форму, сосочків, горбків, щетинок, кущиків, зірчастих лусочок і т. Д. У найпростішому випадку волосок сосочок - короткий округло-конічний виріст клітини шкірки .
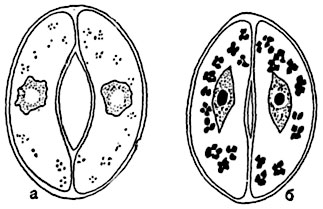
а- продихи відкрито, в замикаючих клітинах дуже дрібні крохмальні зерна, ядро округло-амебоидние обриси, з вакуолей в центрі; б- продихи закрито, в замикаючих клітинах - порівняно великі зерна крохмалю, клітинне ядро веретеноподібно, з великими ядерця в центрі.
Сосочками ж є волоски майже всіх типів в ранній стадії їх утворення. Зустрічаються волоски, що не відділяються перегородкою від здійснила їх клітини шкірки, як наприклад щетинки багатьох бурачнікових. У більшості випадків волосок відділяється перегородкою від здійснила його епідермальній клітини - "материнської клітини волоска". Відчленовують волосок рідко залишається одноклітинним; частіше він стає багатоклітинних. Багатоклітинні волоски дуже різноманітні. Серед них розрізняють 1) лінійні, що складаються з клітин, розташованих в ряд (листя картоплі); 2) кущистий-гіллясті (коров'як, рис. 97, 6 ); 3) лускаті і звездчато-лускаті ( Elaeagnus, Мал. 97, 7, 8 ) І 4) масивні, що представляють як би пучок спаяних ниткоподібних волосків (волоски багатьох губоцвітих). Вміст волосків деяких категорій в дорослому стадії руйнується, порожнини їх зазвичай заповнюються повітрям. Такі волоски, які переважно мають білий, або сіруватий, або жовтуватий колір, можна вважати одним з пристосувань, стримуючих дію на лист або стебло гарячих променів сонця і послаблюють висушує дію вітрів.
Деякі волоски можуть служити рослині захистом від поїдання тваринами; так, тісно розташовані жорсткі, гострі щетинки і повстяний покрив охороняють листя і стебла від об'їдання слимаками і частково від поїдання травоїдними ссавцями.
Пекучі волоски. Пекучий волосок кропиви - велика жива клітиназ бульбашковидні підставою, що сидить в чашоподібної багатоклітинній "підставці". Волосок в мініатюрі як би порожнисту голку шприца і закінчується косо поставленої головкою. Так як оболонка волосини, багатий вапном і кремнеземом, дуже ломка і до того ж в місці переходу "голки" в "головку" дуже тонка, то при щонайменшому дотику тварини або людини до голівці остання відламується, гострий край "голки" встромляє в шкіру, і клітинний сікволоска, що містить пекучі речовини, впорскується в неї (рис. 97, 9-12 ).
Дотик руки до листя деяких Крапов тропічної флори (наприклад, Urtica urentissimaІндонезії) викликає хворобливе або навіть небезпечне для життя запалення.
До покривним тканин належать ще перидерма і кірка (опис їх см. На стор. 173-175).
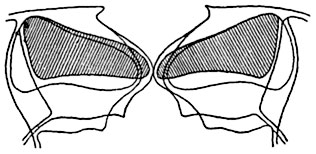
Мал. 96. схематизувати оптичний розріз продихи зимівника ( Heiieborus sp.) В закритому і відкритому стані.
Зовнішні обриси і клітинна порожнину замикаючих клітин показані при закритій щілини продихи і при відкритій щілини. Клітинна порожнину обох замикаючих клітин при закритому продихи заштрихована.
Освітні тканини (меристеми)
Освітні тканини в тілі рослин розташовуються в різних місцях, тому їх ділять на наступні групи (Рис 0; 1).
1. Верхівкове (апікальні) меристемирозташовуються на верхівках, або апексах, осьових органів - стебла, кореня. За допомогою цих меристем вегетативні органи рослин здійснюють своє зростання в довжину.
2. Латеральні меристемихарактерні для осьових органів. Там вони розташовуються концентрично, у вигляді муфти.
3. інтеркалярний,або вставні, меристемипоходять від верхівкових меристем. Це групи клітин, ще не здатних розмножуватися, однак стали на шлях диференціації. Ініціальних клітин серед них немає, але багато спеціалізованих.
4. Ранові меристемизабезпечують відновлення пошкодженої частини тіла. Регенерація починається з дедиференціації, тобто зворотного розвитку від спеціалізованих клітин до меристематические. Вони перетворюються в феллоген,який утворює пробку, Яка покриває поверхню рани. Дедіфференцірованние клітини, ділячись, можуть формувати пухку паренхіматозну тканину - каллус.З нього за певних умов утворюються органи рослин.
покривні тканини
Вони виконують роль прикордонного бар'єру, відокремлюючи нижче лежачі тканини від навколишнього середовища. Первинні покриви рослини складаються тільки з живих клітин. Вторинні і третинні покриви - в основному з мертвих з товстими клітинними стінками.
Основні функції покривних тканин:
· Захист рослини від висихання;
· Захист від потрапляння шкідливих мікроорганізмів;
· Захист від сонячних опіків;
· Захист від механічних пошкоджень;
· Регуляція обміну речовин між рослиною і навколишнім середовищем;
· Сприйняття роздратування.
Первинна покривна тканина - епідерміс, епідерміс . Складається з живих клітин. Утворюється з апікальних меристем. Покриває молоді зростаючі стебла і листя.
Епідерма сформувалася у рослин в зв'язку з виходом з водного середовища проживання на сушу з метою запобігання від висихання. Крім продихів, всі клітини епідерми щільно з'єднані між собою. Зовнішні стіни основних клітин товщі інших. Вся поверхня покрита шаром Кутіна і рослинних восків. Цей шар називається кутикулою(Шкірка). Вона відсутня на зростаючих коренях і підводних частинах рослин. При пересиханні проникність кутикули значно послаблюється.
Крім основних клітин, В епідермісі є і інші, зокрема волоски, або трихоми. Вони бувають одноклітинними і багатоклітинними (Рис.2). Функціонально вони збільшують поверхню епідерми, наприклад, в зоні росту кореня, служити механічним захистом, чіплятися за опору, зменшувати втрати води. Ряд рослин мають залізисті волоски, Наприклад, кропива.
Тільки у вищих рослин в епідермісі є продихи, Які регулюють обмін води і газів. Якщо кутикули немає, то і відсутня потреба в продихах. Устячка - це група клітин, що утворюють устьічнимі апарат,який складається з двох замикаючих клітині прилеглих до них клітин епідерми - побічних клітин. Вони відрізняються від основних епідермальних клітин (рис.3 ). Замикають клітини відрізняються від тих, що оточують їх клітин формою і присутністю великої кількості мітохондрій та нерівномірно потовщеними стінками. Ті, які знаходяться один напроти одного, товщі інших (Рис.4) . Між замикаючими клітинами утворюється устьичная щілину, Яка веде в подустьічное простір, зване подустьічной порожниною.Замикають клітини мають високу фотосинтетичної активністю. У них міститься велика кількість запасного крохмалю і численні мітохондрії.
Число і розподіл устьиц, типи устьічних апаратів широко варіює у різних рослин. Продихи у сучасних мохоподібних відсутні. Фотосинтез у них здійснює гаметофітное покоління, а спорофіти до самостійного існування не здатні.
Зазвичай продихи розташовуються на нижньому боці аркуша. У плаваючих на водній поверхні рослин - на верхній поверхнос. У листя злаків продихи часто розташовуються рівномірно по обидва боки. Такі листи висвітлюються порівняно рівномірно. На 1мм 2 поверхні може розташовуватися від 100 до 700 продихів.
Вторинна покривна тканина (перидерма). Ця тканина приходить на зміну епідермі, коли зелений колір однорічних пагонів змінюється коричневим. Вона багатошарова і складається з центрального шару камбіальних клітин - феллогена.Клітини феллогена, ділячись, назовні відкладають шар феллеми,а всередину - феллодерми(Рис.5).
Феллема, або пробка. Спочатку складається з живих тонкостінних клітин. Згодом їх стінки просочуються суберином і рослинними воском і відмирають. Вміст клітини наповнюється повітрям.
Функції феллеми:
· Запобігає втраті вологи;
· Захищає рослину від механічних пошкоджень;
· Захищає від хвороботворних мікроорганізмів;
· Забезпечує термоізоляцію, так як клітини заповнені повітрям.
Клітини феллогена, розташованого в самій епідермі, що підлягає субепідермальной шарі, рідше - в глибоких шарах первинної кори, є генеруючої основою первинної кори.
Шар пробки не постійний. У ньому відбуваються розриви, які повідомляються з межклетниками, розташованими поруч. При цьому на поверхні утворюються невеликі горбки - чечевички, Які повідомляють простору межклетников з атмосферним повітрям (Рис.6,7).
Восени феллоген під чечевичками відкладає шар опробковевшей клітин, сильно зменшують транспірацію, але не виключає її повністю. Навесні цей шар зсередини руйнується. На світлій корі берези чечевички добре помітні у вигляді темних рисочок.
Третинна покривна тканина (шкірка),так само характерна тільки для деревних форм рослин.
Феллоген багаторазово закладається в більш глибоких шарах кори. Тканини, які виявляються зовні від нього, з часом відмирають, утворюючи кірку. Клітини її мертві і не здатні до розтягування. Однак розташовані глибше живі клітини діляться, що приводить до збільшення поперечного розміру стовбура. Згодом зовнішній шар кірки розривається. Час настання такого розриву є досить постійною величиною для конкретних рослин. У яблуні це відбувається на сьомому році життя, у граба - на п'ятдесятому. У деяких видів не відбувається зовсім. Основна функція кірки - захист від механічних і термічних уражень.